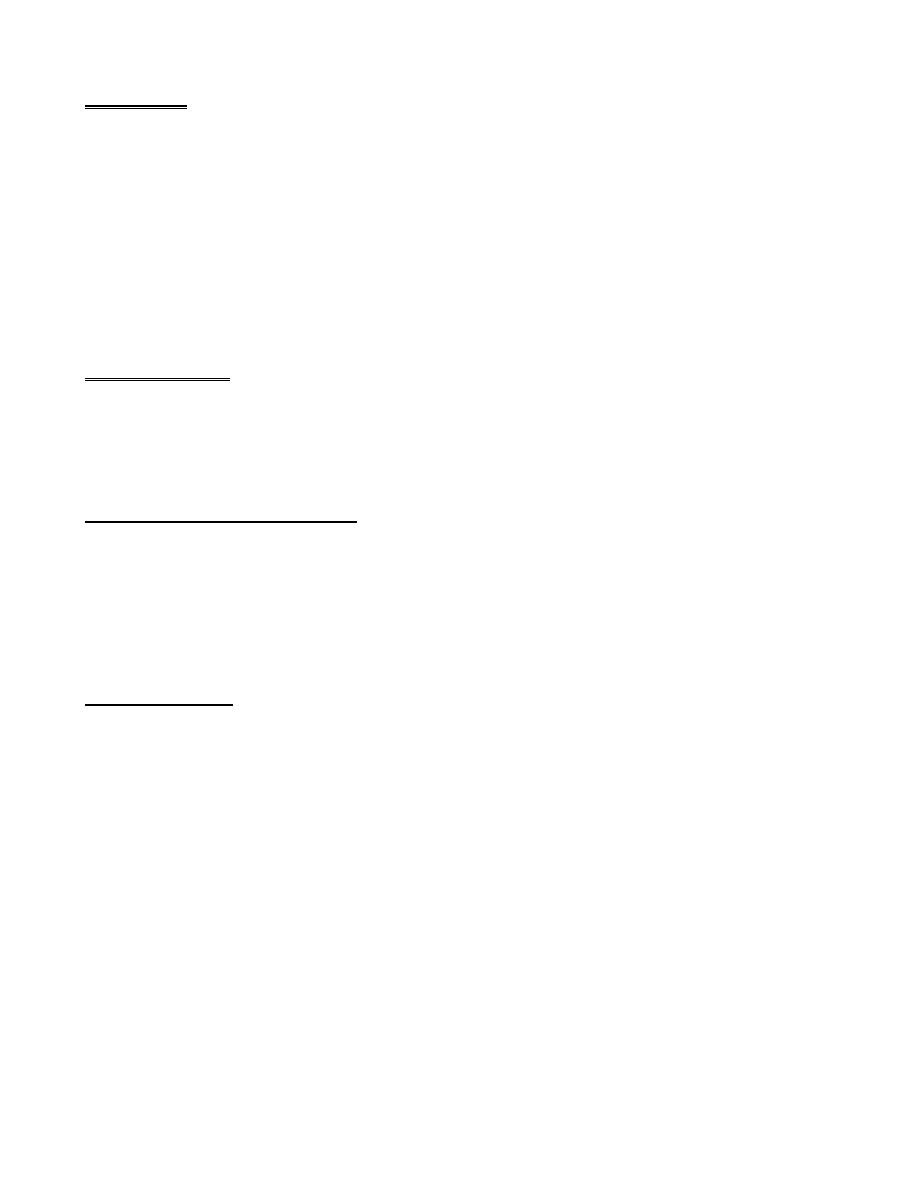
Dr. Affan
Ezzat
Cell Physiology
Lecture one
1
Objectives:
By the end of these two lectures the student will be able to:
Describe the basic components of the cell membrane.
Explain the transport process through cell membranes.
Define diffusion and its types.
List the factors that affect the net rate of diffusion.
Define osmosis.
Explain the active transport of substances through membranes.
Give examples on the types of active transport.
Cell Physiology:
The basic living unit of the body is the cell. Each organ is an aggregate of many different cells held
together by intercellular supporting structures. Each type of cell is specially adapted to perform one or a few
particular functions. For instance, the red blood cells, numbering 25 trillion in each human being, transport
oxygen from the lungs to the tissues. The entire body contains about 100 trillion cells.
Membranous Structures of the Cell
Most organelles of the cell are covered by membranes composed primarily of lipids and proteins. These
membranes include the cell membrane, nuclear membrane, membrane of the endoplasmic reticulum, and
membranes of the mitochondria, lysosomes, and Golgi apparatus. The lipids of the membranes provide a barrier
that impedes the movement of water and water-soluble substances from one cell compartment to another because
water is not soluble in lipids. However, protein molecules in the membrane often do penetrate all the way
through the membrane.
The Cell Membrane:
The cell membrane (also called the plasma membrane), which envelops the cell, is a thin, pliable, elastic
structure only 7.5 to 10 nanometers thick. It is composed almost entirely of proteins and lipids. The approximate
composition is proteins, 55%; phospholipids, 25%; cholesterol, 13%; other lipids, 4%; and carbohydrates, 3%.
Its basic structure is a lipid bilayer, which is a thin, double-layered film of lipids—each layer only one molecule
thick—that is continuous over the entire cell surface. Interspersed in this lipid film are large globular protein
molecules. The basic lipid bilayer is composed of phospholipid molecules. One end of each phospholipid
molecule is soluble in water; that is, it is hydrophilic. The other end is soluble only in fats; that is, it is
hydrophobic. The hydrophilic phosphate portions then constitute the two surfaces of the complete cell
membrane, in contact with intracellular water on the inside of the membrane and extracellular water on the
outside surface. The lipid layer in the middle of the membrane is impermeable to the usual water-soluble
substances, such as ions, glucose, and urea. Conversely, fat-soluble substances, such as oxygen, carbon dioxide,
and alcohol, can penetrate this portion of the membrane with ease.

Dr. Affan
Ezzat
Cell Physiology
Lecture one
2
The Cell Membrane Proteins:
The membrane proteins are globular masses floating in the lipid bilayer, most of which are glycoproteins. Two
types of proteins occur: integral proteins that protrude all the way through the membrane and peripheral
proteins that are attached only to one surface of the membrane and do not penetrate all the way through.
The integral proteins act as (1)structural channels (or pores) through which water molecules and water-soluble
substances, especially ions, can diffuse between the extracellular and intracellular fluids. (2) carrier proteins for
transporting substances that otherwise could not penetrate the lipid bilayer (bind with molecules or ions that are
to be transported; conformational changes in the protein molecules then move the substances through the
interstices of the protein to the other side of the membrane). Both the channel proteins and the carrier proteins
are usually highly selective in the types of molecules or ions that are allowed to cross the membrane.
(3) enzymes (catalyze chemical reactions).
(4) receptors for water-soluble chemicals, such as peptide hormones, that do not easily penetrate the cell
membrane. Interaction of cell membrane receptors with specific ligands that bind to the receptor causes
conformational changes in the receptor protein. This, in turn, enzymatically activates the intracellular part of the
protein or induces interactions between the receptor and proteins in the cytoplasm that act as second messengers,
thereby relaying the signal from the extracellular part of the receptor to the interior of the cell. In this way,
integral proteins spanning the cell membrane provide a means of conveying information about the
environment to the cell interior.
Peripheral protein molecules are often attached to the integral proteins, function almost entirely as (1) enzymes
(2) controllers of transport of substances through the cell membrane “pores.
The Membrane Carbohydrates—The Cell “Glycocalyx.”
Membrane carbohydrates occur in combination with proteins or lipids in the form of glycoproteins or
glycolipids. Most of the integral proteins are glycoproteins, and about one tenth of the membrane lipid molecules
are glycolipids. The “glyco” portions of these molecules protrude to the outside of the cell, dangling outward
from the cell surface. Many other carbohydrate compounds, called proteoglycans—which are mainly
carbohydrate substances bound to small protein cores—are loosely attached to the outer surface of the cell as
well. The entire outside surface of the cell often has a loose carbohydrate coat called the glycocalyx. It has
several important functions:
(1) Have a negative electrical charge, which gives most cells an overall negative surface charge that repels other
negative objects.
(2) The glycocalyx of some cells attaches to the glycocalyx of other cells, thus attaching cells to one another.
(3) Act as receptor substances for binding hormones, such as insulin; when bound, this combination activates
attached internal proteins that, in turn, activate a cascade of intracellular enzymes.
(4) Some carbohydrate types enter into immune reactions.
Dr. Affan
Ezzat
Cell Physiology
Lecture one
3
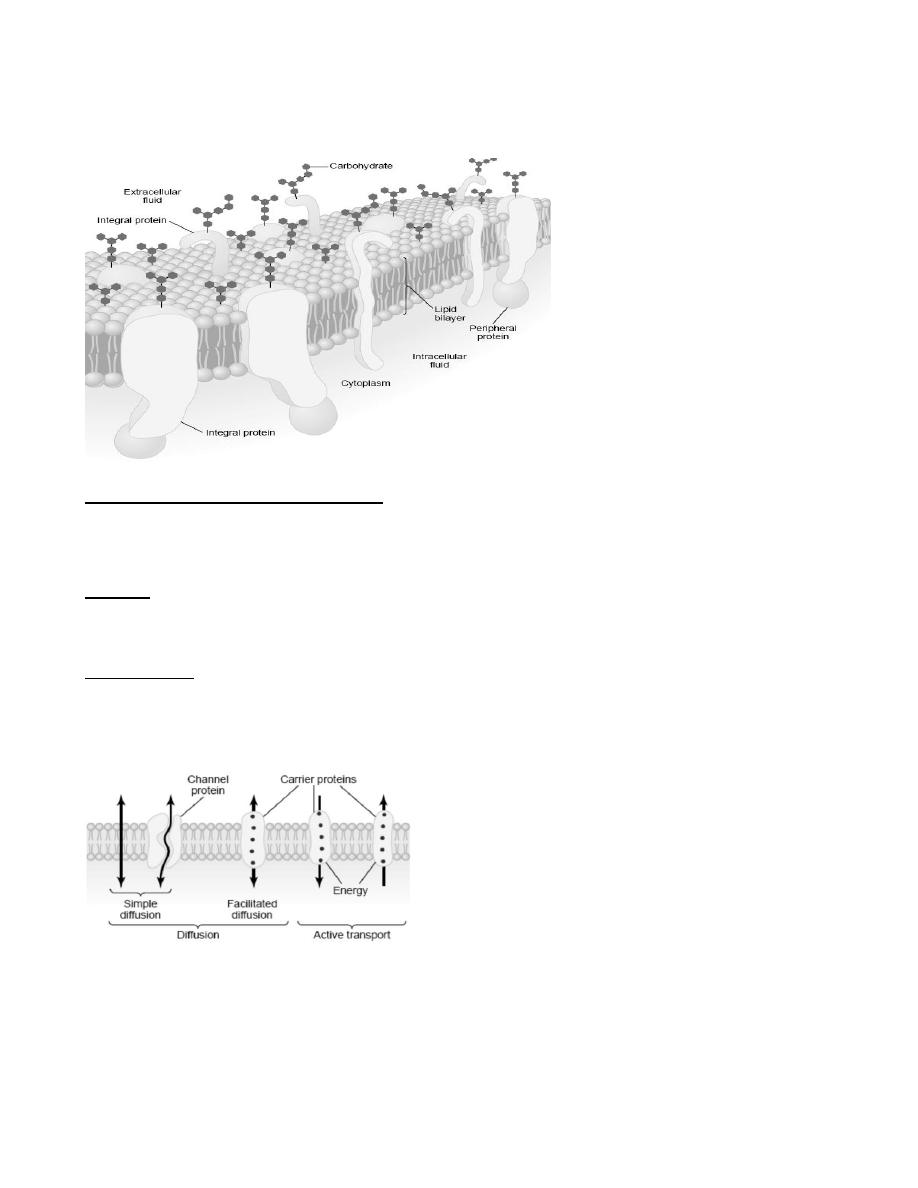
Transport through the cell membrane:
Either directly through the lipid bilayer or through the proteins, occurs by one of two basic processes: diffusion
or active transport.
Diffusion means random molecular movement of substances molecule by molecule, either through
intermolecular spaces in the membrane or in combination with a carrier protein, the energy that causes diffusion
is the energy of the normal kinetic motion of matter.
Active transport means movement of ions or other substances across the membrane in combination with a carrier
protein in such a way that the carrier protein causes the substance to move against an energy gradient, such as
from a low-concentration state to a high-concentration state, It requires an additional source of energy besides
kinetic energy.

Dr. Affan
Ezzat
Cell Physiology
Lecture one
4
Diffusion through the Cell Membrane
Diffusion through the cell membrane is divided into two subtypes called simple diffusion and facilitated
diffusion. Simple diffusion means that kinetic movement of molecules or ions occurs through a membrane
opening or through intermolecular spaces without any interaction with carrier proteins in the membrane. Simple
diffusion can occur through the cell membrane by two pathways: (1) through the interstices of the lipid bilayer if
the diffusing substance is lipid soluble, and (2) through watery channels that penetrate all the way through some
of the large transport proteins.
Facilitated diffusion requires interaction of a carrier protein. The carrier protein aids passage of the molecules or
ions through the membrane by binding chemically with them and shuttling them through the membrane in this
form.
Diffusion of Lipid-Soluble Substances through the Lipid Bilayer.
One of the most important factors that determine how rapidly a substance diffuses through the lipid bilayer is the
lipid solubility of the substance. For instance, the lipid solubilities of oxygen, nitrogen, carbon dioxide, and
alcohols are high, so that all these can dissolve directly in the lipid bilayer and diffuse through the cell membrane
in the same manner that diffusion of water solutes occurs in a watery solution. The rate of diffusion of each of
these substances through the membrane is directly proportional to its lipid solubility. Especially large amounts of
oxygen can be transported in this way; therefore, oxygen can be delivered to the interior of the cell almost as
though the cell membrane did not exist.
Diffusion of Water and Other Lipid-Insoluble Molecules through Protein Channels.
Even though water is highly insoluble in the membrane lipids, it readily passes through channels in protein
molecules that penetrate all the way through the membrane. Other lipid-insoluble molecules can pass through the
protein pore channels in the same way as water molecules if they are water soluble and small enough. However,
as they become larger, their penetration falls off rapidly.
Diffusion Through Protein Channels, and “Gating” of These Channels
Computerized three-dimensional reconstructions of protein channels have demonstrated tubular pathways all the
way from the extracellular to the intracellular fluid. Therefore, substances can move by simple diffusion directly
along these channels from one side of the membrane to the other. The protein channels are distinguished by two
important characteristics: (1) they are often selectively permeable to certain substances, and (2) many of the
channels can be opened or closed by gates.
Selective Permeability of Protein Channels.
Many of the protein channels are highly selective for transport of one or more specific ions or molecules. This
results from the characteristics of the channel itself, such as its diameter, its shape, and the nature of the
electrical charges and chemical bonds along its inside surfaces. To give an example, one of the most important of
the protein channels, the so-called sodium channel, is only 0.3 by 0.5 nanometer in diameter, but more
important, the inner surfaces of this channel are strongly negatively charged, These strong negative charges can
pull small dehydrated sodium ions into these channels, actually pulling the sodium ions away from their
hydrating water molecules. Once in the channel, the sodium ions diffuse in either direction according to the
usual laws of diffusion. Thus, the sodium channel is specifically selective for passage of sodium ions.

Dr. Affan
Ezzat
Cell Physiology
Lecture one
5
Gating of Protein Channels.
Gating of protein channels provides a means of controlling ion permeability of the channels. It is believed that
some of the gates are actual gate like extensions of the transport protein molecule, which can close the opening
of the channel or can be lifted away from the opening by a conformational change in the shape of the protein
molecule itself. The opening and closing of gates are controlled in two principal ways:
1.Voltage gating.
The molecular conformation of the gate or of its chemical bonds responds to the electrical potential across the
cell membrane. when there is a strong negative charge on the inside of the cell membrane, this could cause the
outside sodium gates to remain tightly closed; conversely, when the inside of the membrane loses its negative
charge, these gates would open suddenly and allow tremendous quantities of sodium to pass inward through the
sodium pores. This is the basic mechanism for eliciting action potentials in nerves that are responsible for nerve
signals. The potassium gates are on the intracellular ends of the potassium channels, and they open when the
inside of the cell membrane becomes positively charged. The opening of these gates is partly responsible for
terminating the action potential.
2. Chemical (ligand) gating.
Some protein channel gates are opened by the binding of a chemical substance (a ligand) with the protein; this
causes a conformational or chemical bonding change in the protein molecule that opens or closes the gate. This
is called chemical gating or ligand gating. One of the most important instances of chemical gating is the effect of
acetylcholine on the so-called acetylcholine channel. Acetylcholine opens the gate of this channel, providing a
negatively charged pore about 0.65 nanometer in diameter that allows uncharged molecules or positive ions
smaller than this diameter to pass through. This gate is important for the transmission of nerve signals from one
nerve cell to another and from nerve cells to muscle cells to cause muscle contraction.
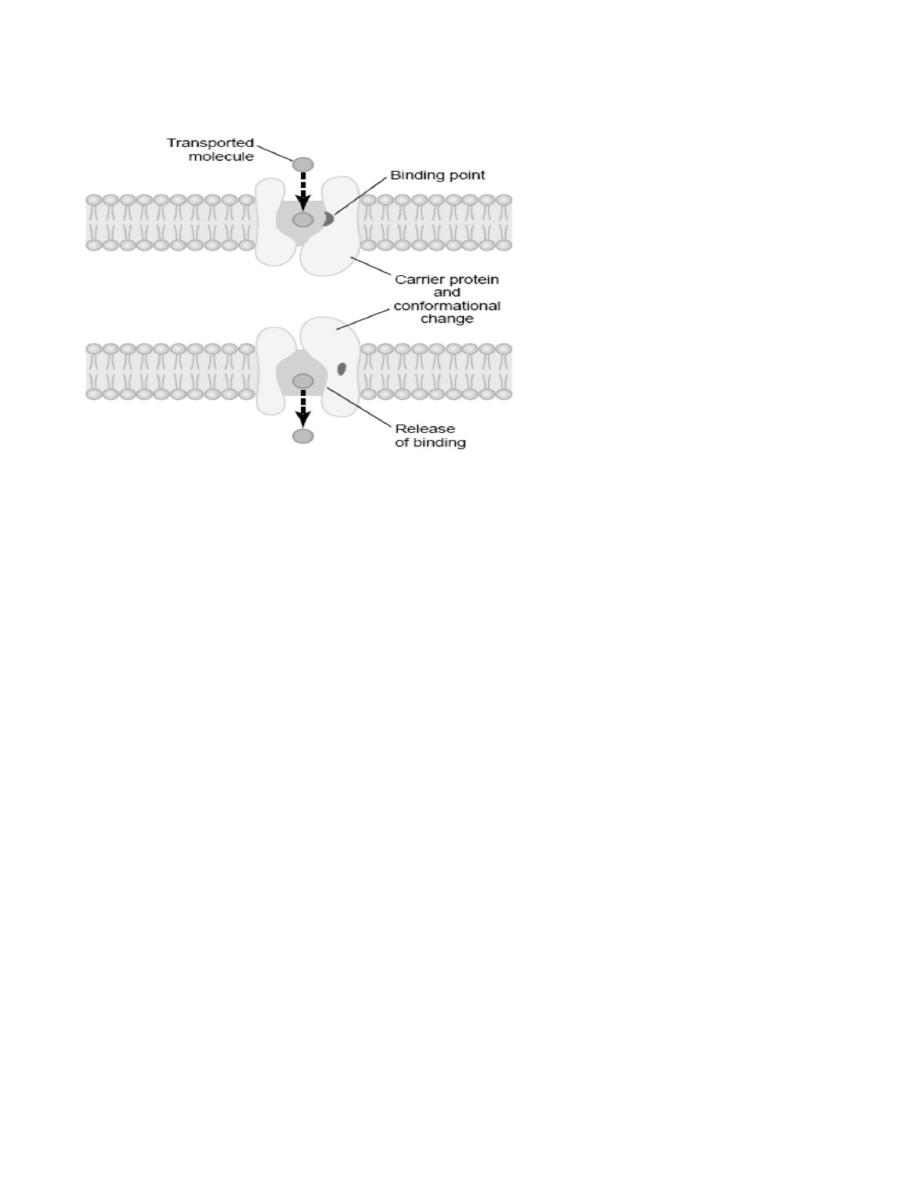
Facilitated Diffusion
Facilitated diffusion is also called carrier-mediated diffusion because a substance transported in this manner
diffuses through the membrane using a specific carrier protein to help. That is, the carrier facilitates diffusion of
the substance to the other side.
Facilitated diffusion differs from simple diffusion in the following important way: Although the rate of simple
diffusion through an open channel increases proportionately with the concentration of the diffusing substance, in
facilitated diffusion the rate of diffusion approaches a maximum, called Vmax, as the concentration of the
diffusing substance increases, the rate of simple diffusion continues to increase proportionately, but in facilitated
diffusion, the rate of diffusion cannot rise greater than the Vmax level. What is it that limits the rate of
facilitated diffusion?
a carrier protein with a pore large enough to transport a specific molecule partway through. It also shows a
binding “receptor” on the inside of the protein carrier. The molecule to be transported enters the pore and
becomes bound. Then, in a fraction of a second, a conformational or chemical change occurs in the carrier
protein, so that the pore now opens to the opposite side of the membrane. Because the binding force of the
receptor is weak, the thermal motion of the attached molecule causes it to break away and to be released on the
opposite side of the membrane. The rate at which molecules can be transported by this mechanism can never be
greater than the rate at which the carrier protein molecule can undergo change back and forth between its two
states.
Dr. Affan
Ezzat
Cell Physiology
Lecture one
6