
1
The Autonomic Nervous System (ANS):
After studying these lectures, you should be able to. .
1. Compare the structures and pathways of the autonomic system with
those involved in the control of skeletal muscle.
2. Explain how autonomic innervation of involuntary effectors differs
from the innervation of skeletal muscle.
3. Describe the structure and general functions of the sympathetic
division of the autonomic system.
4. Describe the structure and general functions of the parasympathetic
division of the autonomic system.
5. List the neurotransmitters of the preganglionic and postganglionic
neurons of the sympathetic and parasympathetic systems.
6. Describe the structural and functional relationships between the
sympathetic system and the adrenal medulla.
7. Distinguish between the different types of adrenergic receptors and
explain the physiological and clinical significance of these receptors.
8. Explain how the autonomic system is controlled by the brain.
The ANS coordinates cardiovascular, respiratory, digestive, urinary and
reproductive functions.
This system helps to control arterial pressure, gastrointestinal motility,
gastrointestinal secretions, urinary bladder, sweating, body temperature, and
many other activities. Some of theses activity regulated partially and some
others entirely regulated by ANS.
The most striking characteristic of ANS is the rapidity and intensity with
which it can change visceral functions. For example, it can increase heart rate
twice normal within 3 to 5 seconds, and within 10 to 15 seconds the arterial
pressure can be double; or at other extreme, the arterial pressure can
decrease within 5 seconds to fainting!
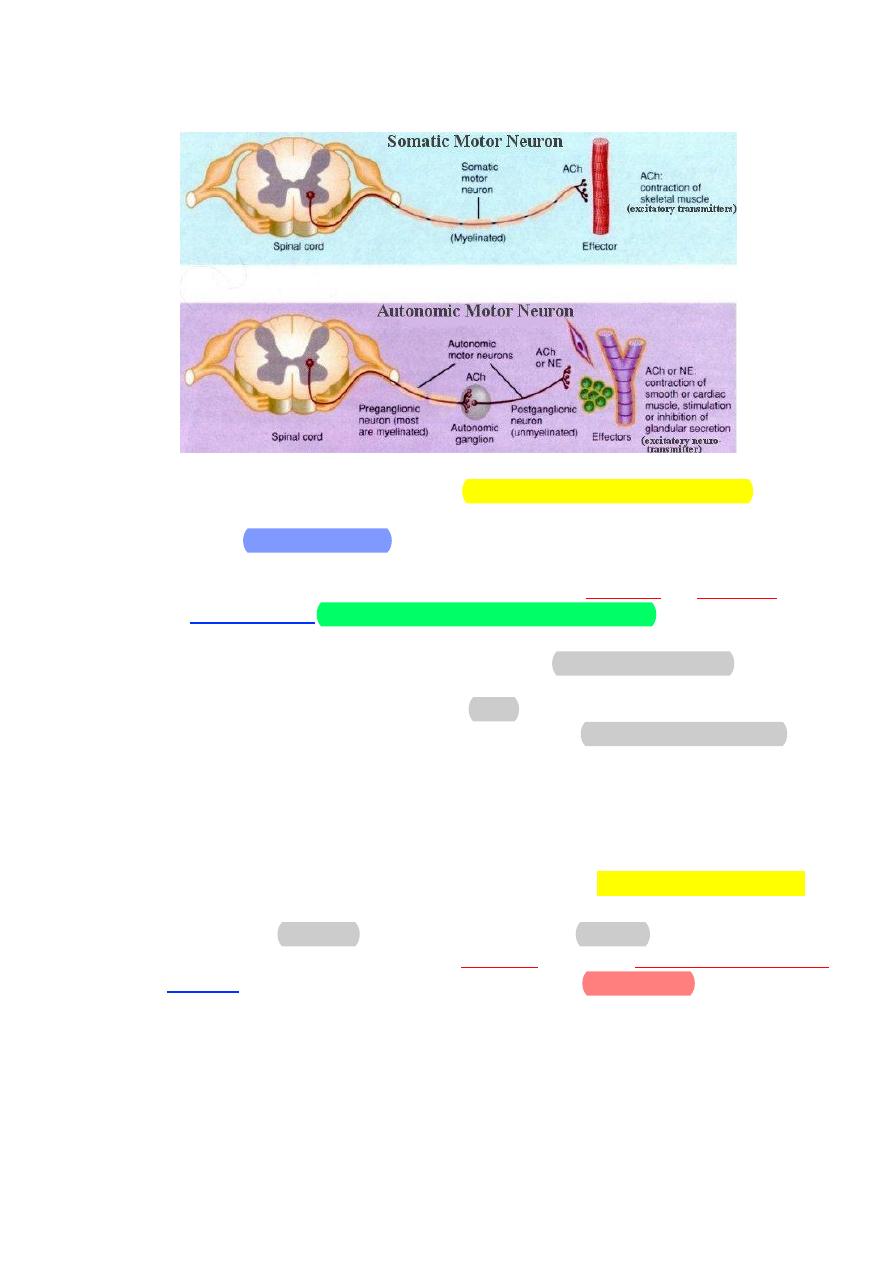
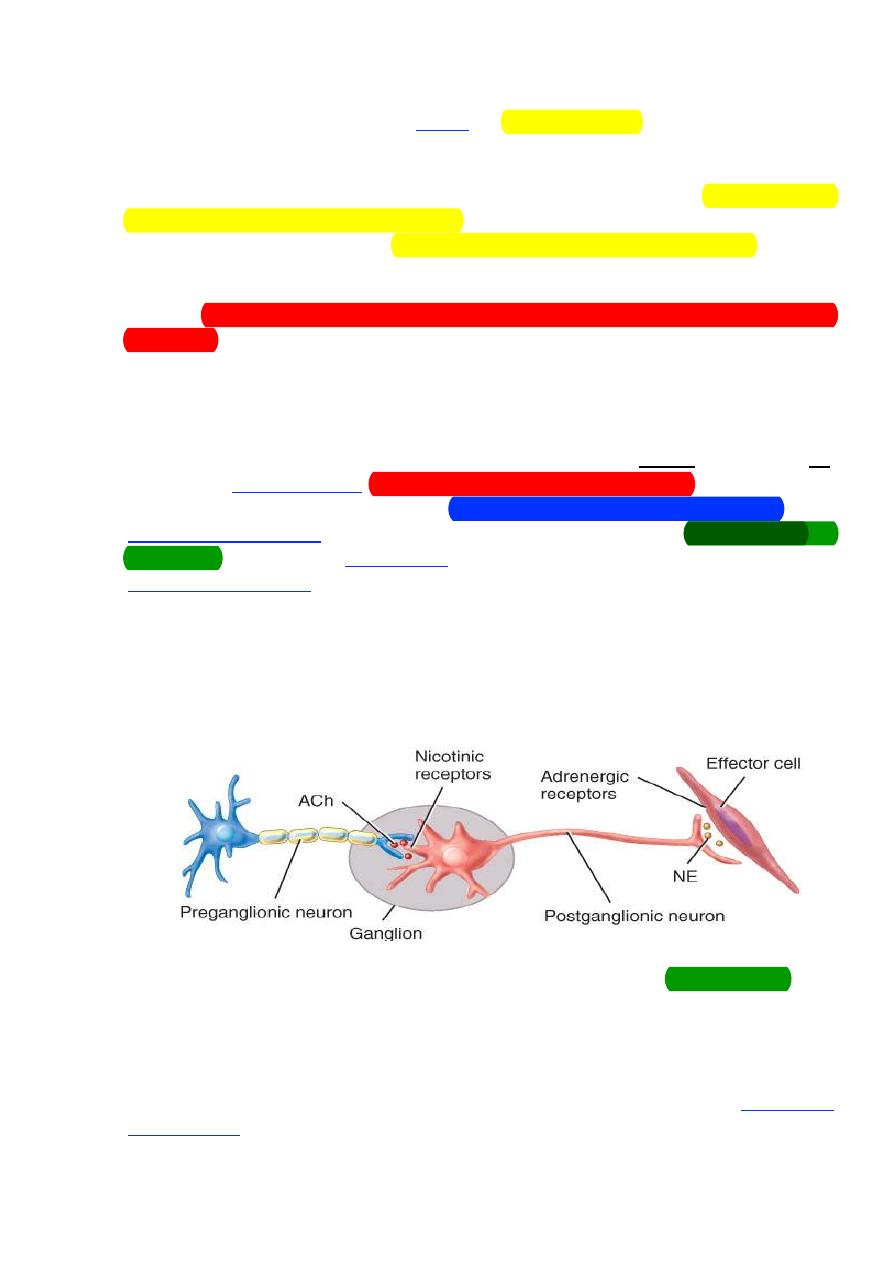
Basic anatomy of ANS:
• Preganglionic neuron
– Cell body in brain or spinal cord.
– Axon is myelinated type B fiber that extends to autonomic ganglion.
• Postganglionic neuron
– Cell body lies outside the CNS in an autonomic ganglion
–
Axon is unmyelinated type C fiber that terminates in a visceral
effector.
2
The ANS is composed of 2 anatomically and functionally distinct
divisions, the sympathetic system and the parasympathetic system. Both
systems are tonically active. In other words, they provide some degree of
nervous input to a given tissue at all times. Therefore, the frequency of
discharge of neurons in both systems can either increase or decrease. As a
result, tissue activity may be either enhanced or inhibited. This characteristic
of the ANS improves its ability to more precisely regulate a tissue's function.
Without tonic activity, nervous input to a tissue could only increase.
Many tissues are innervated by both systems. Because the sympathetic
system and the parasympathetic system typically have opposing effects on a
given tissue, increasing the activity of one system while simultaneously
decreasing the activity of the other results in very rapid and precise control of
a tissue's function.
Sympathetic Division
The sympathetic division is also called the thoracolumbar division of
the autonomic system because its preganglionic fibers exit the spinal cord
from the first thoracic (T1) to the second lumbar (L2) levels. Most
sympathetic nerve fibers, however, separate from the somatic motor fibers
and synapse with postganglionic neurons within a double row of sympathetic
ganglia, called paravertebral ganglia, located on either side of the spinal
cord. Ganglia within each row are interconnected, forming a sympathetic
chain of ganglia that parallels the spinal cord on each lateral side.
3
The myelinated preganglionic sympathetic axons exit the spinal cord in
the ventral roots of spinal nerves, but they soon diverge from the spinal
nerves within short pathways. The axons within each ramus enter the
sympathetic chain of ganglia, where they can travel to ganglia at different
levels and synapse with postganglionic sympathetic neurons.
These ganglion chains, which run parallel immediately along either side
of the spinal cord, each consist of 22 ganglia. The preganglionic neuron may
exit the spinal cord and synapse with a postganglionic neuron in a ganglion at
the same spinal cord level from which it arises. The preganglionic neuron may
also travel more upward or downward in the ganglion chain to synapse with

4
postganglionic neurons in ganglia at other levels. In fact, a single
preganglionic neuron may synapse with several postganglionic neurons in
many different ganglia.
The long postganglionic neurons originating in the ganglion chain then
travel outward and terminate on the effector tissues. This divergence of the
preganglionic neuron results in coordinated sympathetic stimulation to
tissues throughout the body. The concurrent stimulation of many organs and
tissues in the body is referred to as a mass sympathetic discharge.
Other preganglionic neurons exit the spinal cord and pass through the
ganglion chain without synapsing with a postganglionic neuron. Instead, the
axons of these neurons travel more peripherally and synapse with
postganglionic neurons in one of the sympathetic collateral ganglia.
These include the celiac, superior mesenteric and inferior mesenteric
ganglia. Postganglionic fibers that arise from the collateral ganglia innervate
organs of the digestive, urinary, and reproductive systems.
Finally, the preganglionic neuron may travel to the adrenal medulla and
synapse directly with this glandular tissue. The cells of the adrenal medulla
have the same embryonic origin as neural tissue and, in fact, function as
modified postganglionic neurons. Instead of the release of neurotransmitter
directly at the synapse with an effector tissue, the secretory products of the
adrenal medulla are picked up by the blood and travel throughout the body to
all of the effector tissues of the sympathetic system.
An important feature of this system, which is quite distinct from the
parasympathetic system, is that the postganglionic neurons of the
sympathetic system travel within each of the 31 pairs of spinal nerves. This
allows for the distribution of sympathetic nerve fibers to the effectors of the
skin including blood vessels and sweat glands. In fact, most innervated blood
vessels in the entire body, primarily arterioles and veins, receive only
sympathetic nerve fibers. Therefore, vascular smooth muscle tone and
sweating are regulated by the sympathetic system only. In addition, the
sympathetic system innervates structures of the head (eye, salivary glands,
and mucus membranes of the nasal cavity), thoracic viscera (heart, lungs) and
viscera of the abdominal and pelvic cavities (eg, stomach, intestines, pancreas,
spleen, adrenal medulla, and urinary bladder).
Parasympathetic Division
The preganglionic neurons of the parasympathetic system arise from
several nuclei of the brainstem and from the sacral region of the spinal cord
(segments S2-S4). The axons of the preganglionic neurons are quite long
compared to those of the sympathetic system and synapse with
postganglionic neurons within terminal ganglia which are close to or
embedded within the effector tissues. The axons of the postganglionic
5
neurons, which are very short, then provide input to the cells of that effector
tissue.
The preganglionic neurons that arise from the brainstem exit the CNS
through the cranial nerves. The occulomotor nerve (III) innervates the eyes;
the facial nerve (VII) innervates the lacrimal gland, the salivary glands and
the mucus membranes of the nasal cavity; the glossopharyngeal nerve (IX)
innervates the parotid (salivary) gland; and the vagus nerve (X) innervates
the viscera of the thorax and the abdomen (e.g., heart, lungs, stomach,
pancreas, small intestine, upper half of the large intestine, and liver). The
physiological significance of this nerve in terms of the influence of the
parasympathetic system is clearly illustrated by its widespread distribution
and the fact that 75% of all parasympathetic fibers are in the vagus nerve. The
preganglionic neurons that arise from the sacral region of the spinal cord exit
the CNS and join together to form the pelvic nerves. These nerves innervate
the viscera of the pelvic cavity (eg, lower half of the large intestine and organs
of the renal and reproductive systems).
Because the terminal ganglia are located within the innervated tissue,
there is typically little divergence in the parasympathetic system compared to
the sympathetic system. In many organs, there is a 1:1 ratio of preganglionic
fibers to postganglionic fibers. Therefore, the effects of the parasympathetic
system tend to be more discrete and localized, with only specific tissues being
stimulated at any given moment, compared to the sympathetic system where
a more diffuse discharge.
Neurotransmitters of the Autonomic Nervous
System:
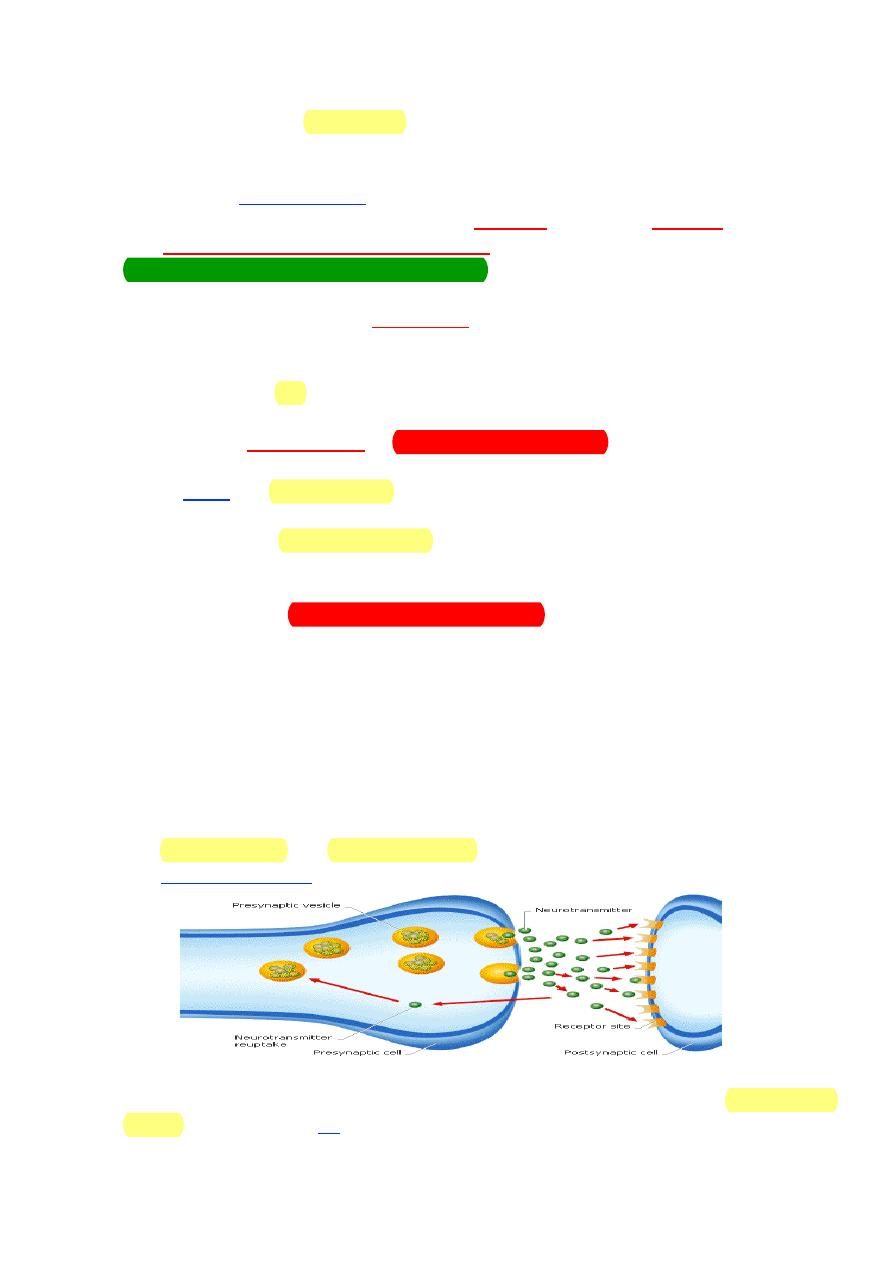
The 2 most common neurotransmitters released by neurons of the ANS
are acetylcholine and norepinephrine. Neurotransmitters are synthesized in
the axon varicosities and stored in vesicles for subsequent release.
Nerve fibers that release acetylcholine are referred to as cholinergic
fibers. These include all preganglionic fibers of the ANS, both sympathetic and
37910
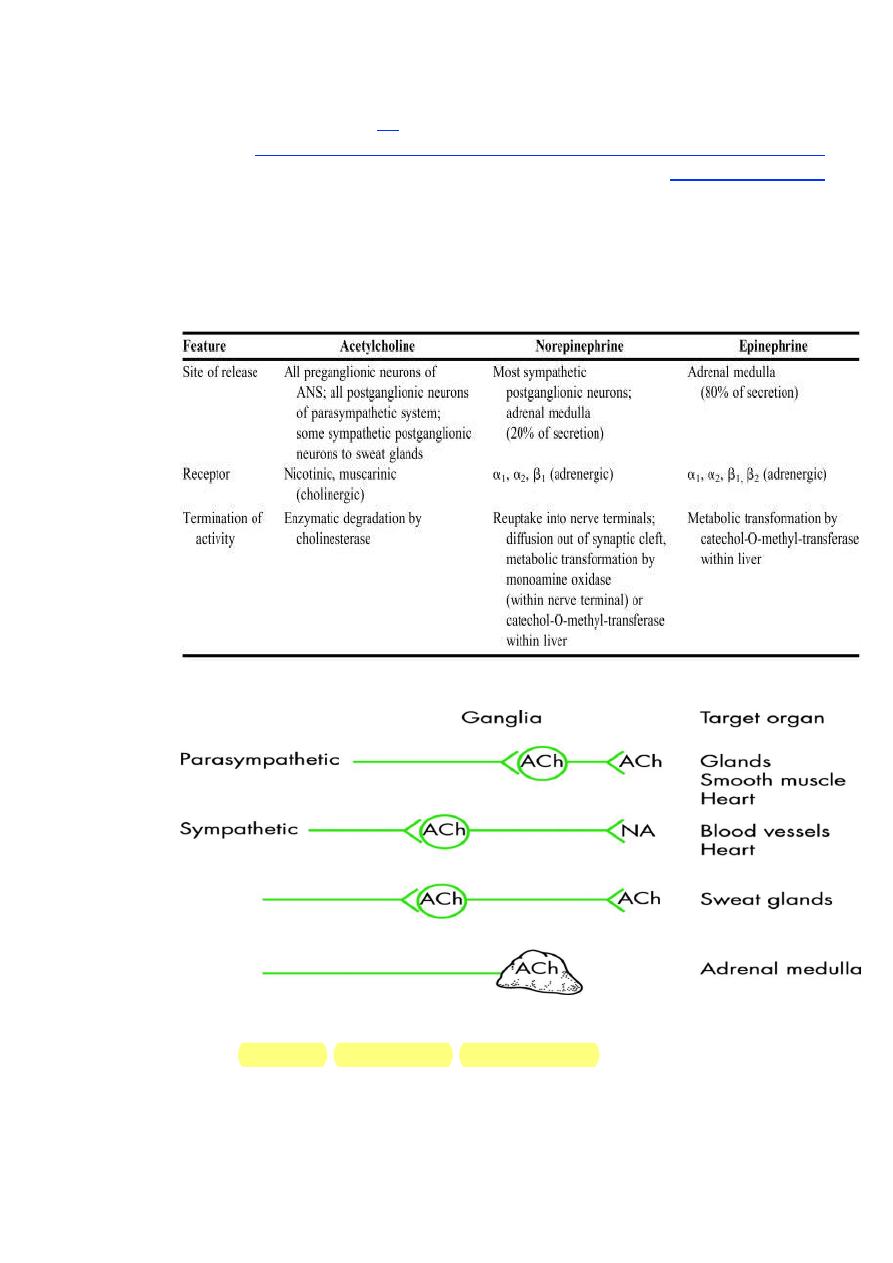
6
parasympathetic systems; all postganglionic fibers of the parasympathetic
system; and sympathetic postganglionic fibers innervating sweat glands.
Nerve fibers that release norepinephrine are referred to as adrenergic fibers.
Most sympathetic postganglionic fibers release norepinephrine.
Distinguishing Features of Neurotransmitters of the Autonomic
Nervous System were summarized in this table.
As previously mentioned, the cells of the adrenal medulla are
considered modified sympathetic postganglionic neurons. Instead of a
neurotransmitter, these cells release hormones into the blood. Approximately
20% of the hormonal output of the adrenal medulla is norepinephrine. The
remaining 80% is epinephrine. Unlike true postganglionic neurons in the
7
sympathetic system, the adrenal medulla contains an enzyme that methylates
norepinephrine to form epinephrine. The synthesis of epinephrine, also
known as adrenaline, is enhanced under conditions of stress. These 2
hormones released by the adrenal medulla are collectively referred to as the
catecholamines.
Receptors for Autonomic Neurotransmitters
As discussed in the previous section, all of the effects of the ANS in
tissues and organs throughout the body, including smooth muscle contraction
or relaxation, alteration of myocardial activity, and increased or decreased
glandular secretion, are carried out by only 3 substances, acetylcholine,
norepinephrine, and epinephrine. Furthermore, each of these substances may
stimulate activity in some tissues and inhibit activity in others.
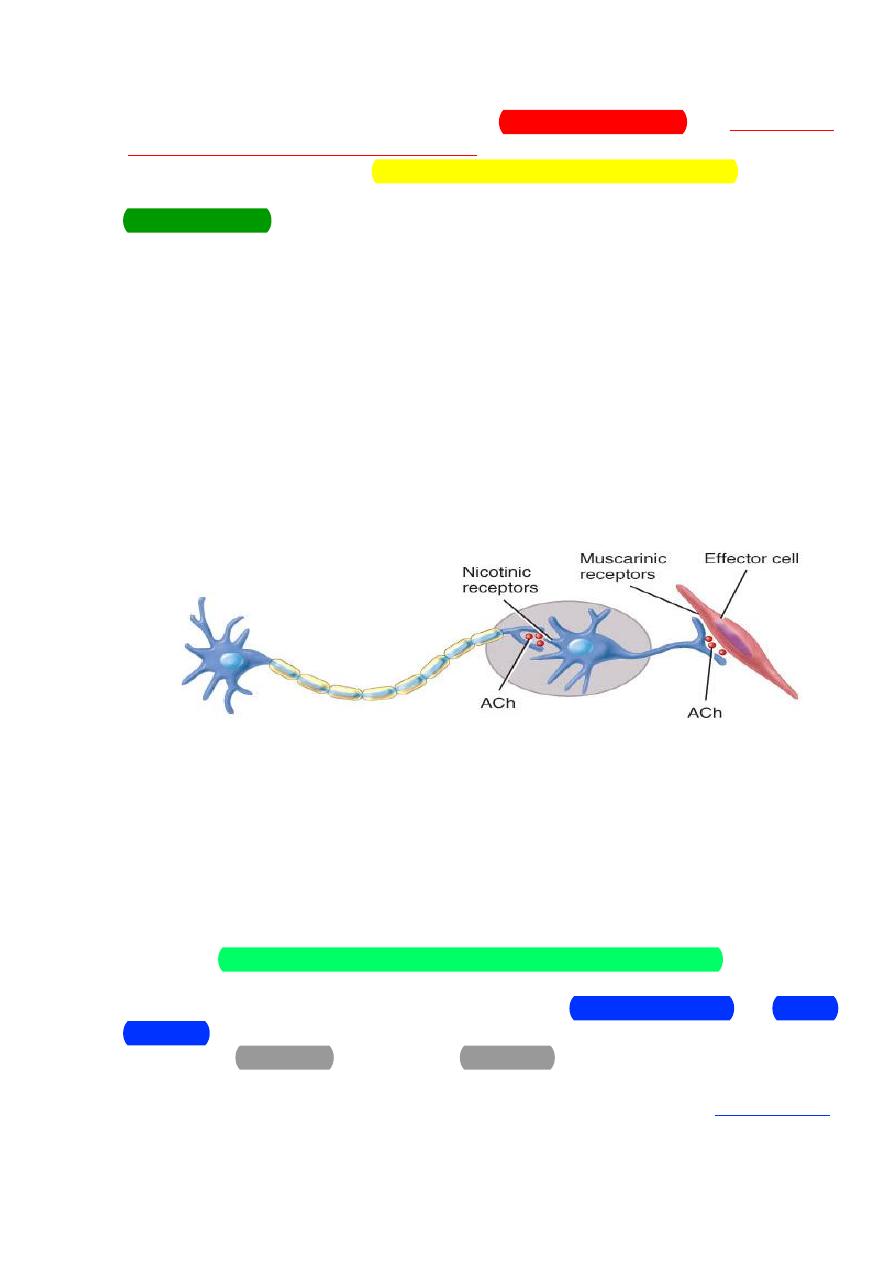
The cholinergic nerve fibers:
Cholinergic Neurons
• Cholinergic neurons release the neurotransmitter
In the ANS, the cholinergic neurons include:
1) All sympathetic and parasympathetic preganglionic neurons
2) Sympathetic postganglionic neurons that innervate most sweat
glands
3) All parasympathetic postganglionic neurons
Acetylcholine is stored in synaptic vesicles and released by exocytosis.
It diffuses across the synaptic cleft and binds with specific cholinergic
receptors, integral proteins in the postsynaptic plasma membrane.
●
Excitation or inhibition depending upon receptor subtype and organ
involved
●
The two types of cholinergic receptors are nicotinic and muscarinic
receptors.
●
Activation of nicotinic receptors causes excitation of the postsynaptic
cell.
=norepinephrine & epinephrine
8
●
Nicotinic receptors are found on the cell bodies of all postganglionic
neurons, both sympathetic and parasympathetic, in the ganglia of the ANS
(and at neuromuscular junction). Acetylcholine released from the
preganglionic neurons binds to these nicotinic receptors and causes a rapid
increase in the cellular permeability to Na
+
ions and Ca
++
ions. The resulting
influx of these 2 cations causes depolarization and excitation of the
postganglionic neurons the ANS pathways.
●
Nicotine mimics the action of acetylcholine by binding to these
receptors.
●
Muscarinic receptors are found on the cell membranes of the effector
tissues and are linked to G proteins and second messenger systems which
carry out the intracellular effects.
●
Activation of muscarinic receptors can cause either excitation or
inhibition depending on the cell that bears the receptors. For example,
muscarinic receptor stimulation in the myocardium is inhibitory and
decreases heart rate while stimulation of these receptors in the lungs is
excitatory, causing contraction of airway smooth muscle and
bronchoconstriction.
●
Muscarinic receptors are found on plasma membranes of all
parasympathetic effectors. Examples: smooth muscle, cardiac muscle and
glands.
The adrenergic nerve fibers:
In
The ANS, adrenergic neurons release norepinephrine (noradrenalin).
●
Most sympathetic postganglionic neurons are adrenergic.
NE is synthesized and stored in synaptic vesicles and released by
exocytosis.
●
Molecules of NE diffuse across the synaptic cleft and bind to specific
adrenergic receptors on the postsynaptic membrane, causing either excitation
or inhibition of the effector cell.

9
●
The main types of adrenergic receptors are alpha and beta receptors.
These receptors are found on visceral effectors innervated by most
sympathetic postganglionic axons.
These receptors are further classified into subtypes.
– Alpha1 and Beta1 receptors produce excitation
– Alpha2 and Beta2 receptors cause inhibition
●
Effects triggered by adrenergic neurons typically are longer lasting
than those triggered by cholinergic neurons
●
Cells of most effectors contain either alpha or beta receptors.
Norepinephrine stimulates alpha receptors more strongly than beta receptors
●
All of these receptors are linked to G proteins and second messenger
systems which carry out the intracellular effects.
Alpha receptors are the more abundant of the adrenergic receptors. Of
the 2 subtypes, α
1
receptors are more widely distributed on the effector
tissues. Alpha one receptor stimulation leads to an increase in intracellular
calcium. As a result, these receptors tend to be excitatory. For example,
stimulation of α
1
receptors causes contraction of vascular smooth muscle
resulting in vasoconstriction and increased glandular secretion by way of
exocytosis.
Termination of Neurotransmitter Activity
For any substance to serve effectively as a neurotransmitter, it must be
rapidly inactivated or removed from the synapse or, in this case, the
neuroeffector junction. This is necessary in order to allow new signals to get
through and influence effector tissue function.
●
The primary mechanism used by cholinergic synapses is enzymatic
degradation. Acetylcholinesterase hydrolyzes acetylcholine to its component
choline and acetate. It is one of the fastest acting enzymes in the body and
acetylcholine removal occurs in less than 1 msec.
●
The most important mechanism for the removal of norepinephrine
from the neuroeffector junction is the reuptake of this neurotransmitter into
the sympathetic nerve that released it. Norepinephrine may then be
metabolized intraneuronally by monoamine oxidase (MAO).
●
The circulating catecholamines, epinephrine and norepinephrine, are
inactivated by catechol-O-methyltransferase (COMT) in the liver.

11
● Certain postganglionic autonomic axons produce their effects
through mechanisms that do not involve either norepinephrine or
acetylcholine. This can be demonstrated experimentally by the inability of
drugs that block adrenergic and cholinergic effects from inhibiting the
actions of those autonomic axons.
These axons, consequently, have been termed “nonadrenergic,
noncholinergic fibers.” Proposed neurotransmitters for these axons include
ATP, a polypeptide called vasoactive intestinal peptide (VIP), and nitric oxide
(NO).
The nonadrenergic, noncholinergic parasympathetic axons that
innervate the blood vessels of the penis cause the smooth muscles of these
vessels to relax, thereby producing vasodilation and a consequent erection of
the penis.
In a similar manner, nitric oxide appears to function as the autonomic
neurotransmitter that causes vasodilation of cerebral arteries.
Studies suggest that nitric oxide is not stored in synaptic vesicles, as are
other neurotransmitters, but instead is produced immediately when Ca2+
enters the axon terminal in response to action potentials. This Ca2+
indirectly activates nitric oxide synthetase, the enzyme that forms nitric
oxide from the amino acid L-arginine. Nitric oxide then diffuses across the
synaptic cleft and promotes relaxation of the postsynaptic smooth muscle
cells.
Nitric oxide can produce relaxation of smooth muscles in many
organs, including the stomach, small intestine, large intestine, and urinary
bladder.
Indeed, nitric oxide is a member of a class of local tissue regulatory
molecules called paracrine regulators. Regulation can therefore be a complex
process involving the interacting effects of different neurotransmitters,
hormones, and paracrine regulators.
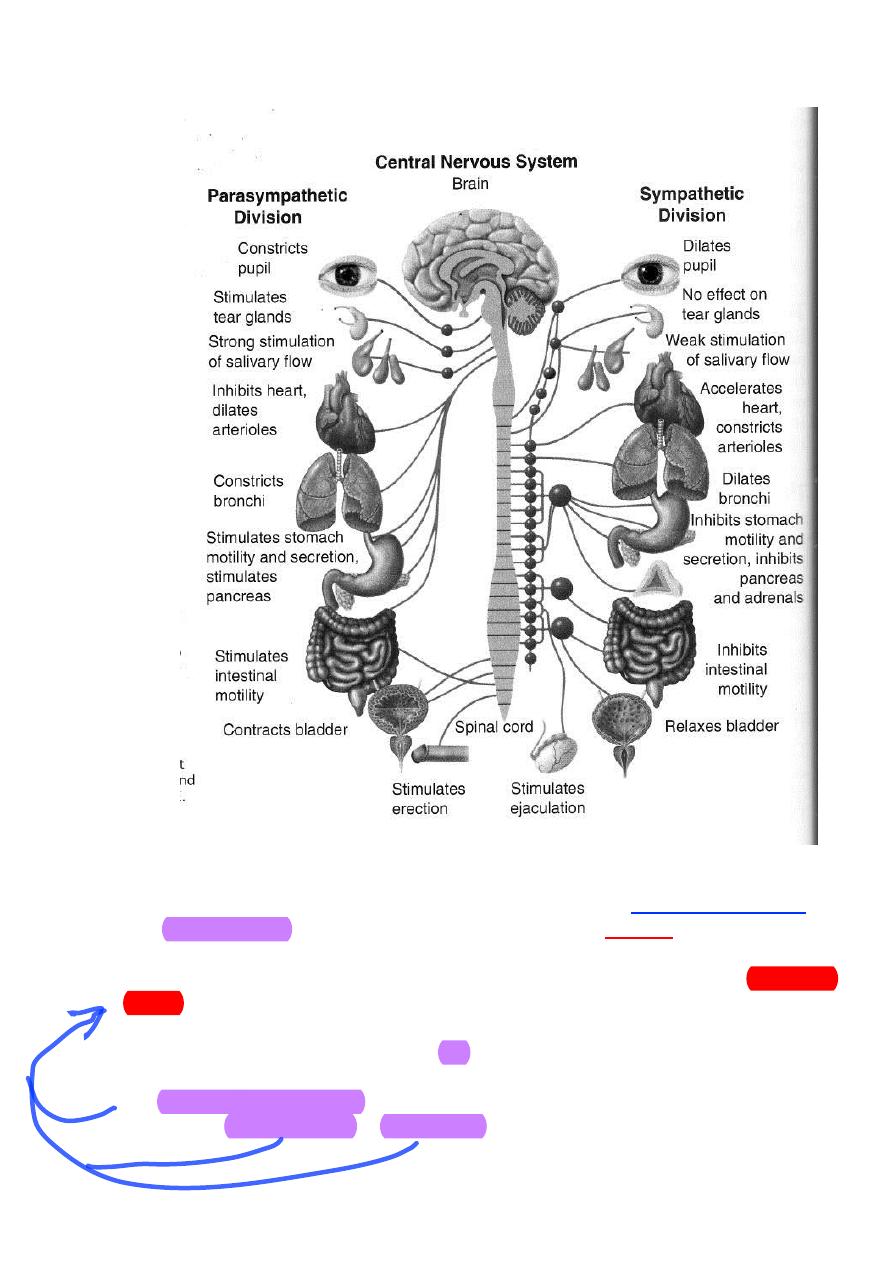
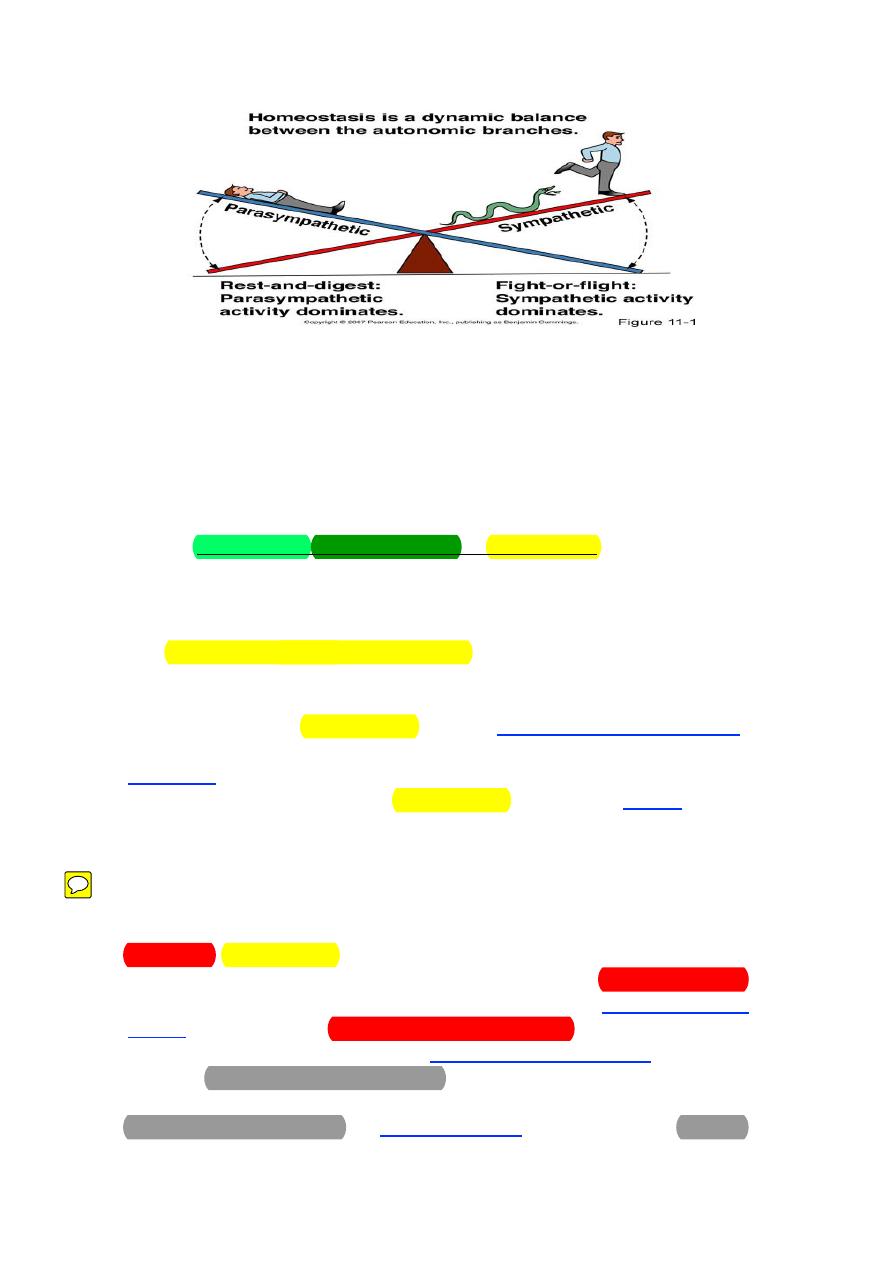
Functions of the Autonomic Nervous System
The 2 divisions of the ANS are dominant under different conditions. As
stated previously, the sympathetic system is activated during emergency
“fight-or-flight” reactions and during exercise. The parasympathetic system is
predominant during quiet conditions (“rest and digest”). As such, the
physiological effects caused by each system are quite predictable. In other
words, all of the changes in organ and tissue function induced by the
sympathetic system work together to support strenuous physical activity and
the changes induced by the parasympathetic system are appropriate for when
the body is resting.

11

12
The “fight-or-flight” reaction elicited by the sympathetic system is
essentially a whole body response.
Changes in organ and tissue function throughout the body are
coordinated so that there is an increase in the delivery of well-oxygenated,
nutrient-rich blood to the working skeletal muscles. Both heart rate and
myocardial contractility are increased so that the heart pumps more blood per
minute. Sympathetic stimulation of vascular smooth muscle causes
widespread vasoconstriction, particularly in the organs of the gastrointestinal
system and in the kidneys. This vasoconstriction serves to “redirect” or
redistribute the blood away from these metabolically inactive tissues and
toward the contracting muscles. Bronchodilation in the lungs facilitates the
movement of air in and out of the lungs so that the uptake of oxygen from the
atmosphere and the elimination of carbon dioxide from the body are
maximized. An enhanced rate of glycogenolysis (breakdown of glycogen into
its component glucose molecules) and gluconeogenesis (formation of new
glucose from noncarbohydrate sources) in the liver increases the
concentration of glucose molecules in the blood. This is necessary for the
brain as glucose is the only nutrient molecule that it can utilize to form
metabolic energy. An enhanced rate of lipolysis in adipose tissue increases the
concentration of fatty acid molecules in the blood. Skeletal muscles then
utilize these fatty acids to form metabolic energy for contraction. Generalized
sweating elicited by the sympathetic system enables the individual to
thermoregulate during these conditions of increased physical activity and
heat production. Finally, the eye is adjusted such that the pupil dilates letting
more light in toward the retina (mydriasis) and the lens adapts for distance
vision.

13
The parasympathetic system decreases heart rate which helps to
conserve energy under resting conditions.
Salivary secretion is enhanced to facilitate the swallowing of food.
Gastric motility and secretion are stimulated to begin the processing of
ingested food. Intestinal motility and secretion are also stimulated to continue
the processing and to facilitate the absorption of these nutrients. Both
exocrine and endocrine secretion from the pancreas is promoted. Enzymes
released from the exocrine glands of the pancreas contribute to the chemical
breakdown of the food in the intestine and insulin released from the
pancreatic islets promotes the storage of nutrient molecules within the tissues
once they are absorbed into the body. Another bodily maintenance type of
function caused by the parasympathetic system is contraction of the urinary
bladder which results in urination. Finally, the eye is adjusted such that the
pupil contracts (miosis) and the lens adapts for near vision.
Adrenal Medulla
A mass sympathetic discharge, which typically occurs during the “fight-
or-flight” response and during exercise, involves the simultaneous stimulation
of organs and tissues throughout the body. Included among these tissues are
the adrenal medullae which release epinephrine (80%) and norepinephrine
(20%) into the blood. In large part, the indirect effects of these
catecholamines are similar to and, therefore, reinforce those of direct
sympathetic stimulation. However, there are some important differences in
the effects of the circulating catecholamines and those of norepinephrine
released from sympathetic nerves.
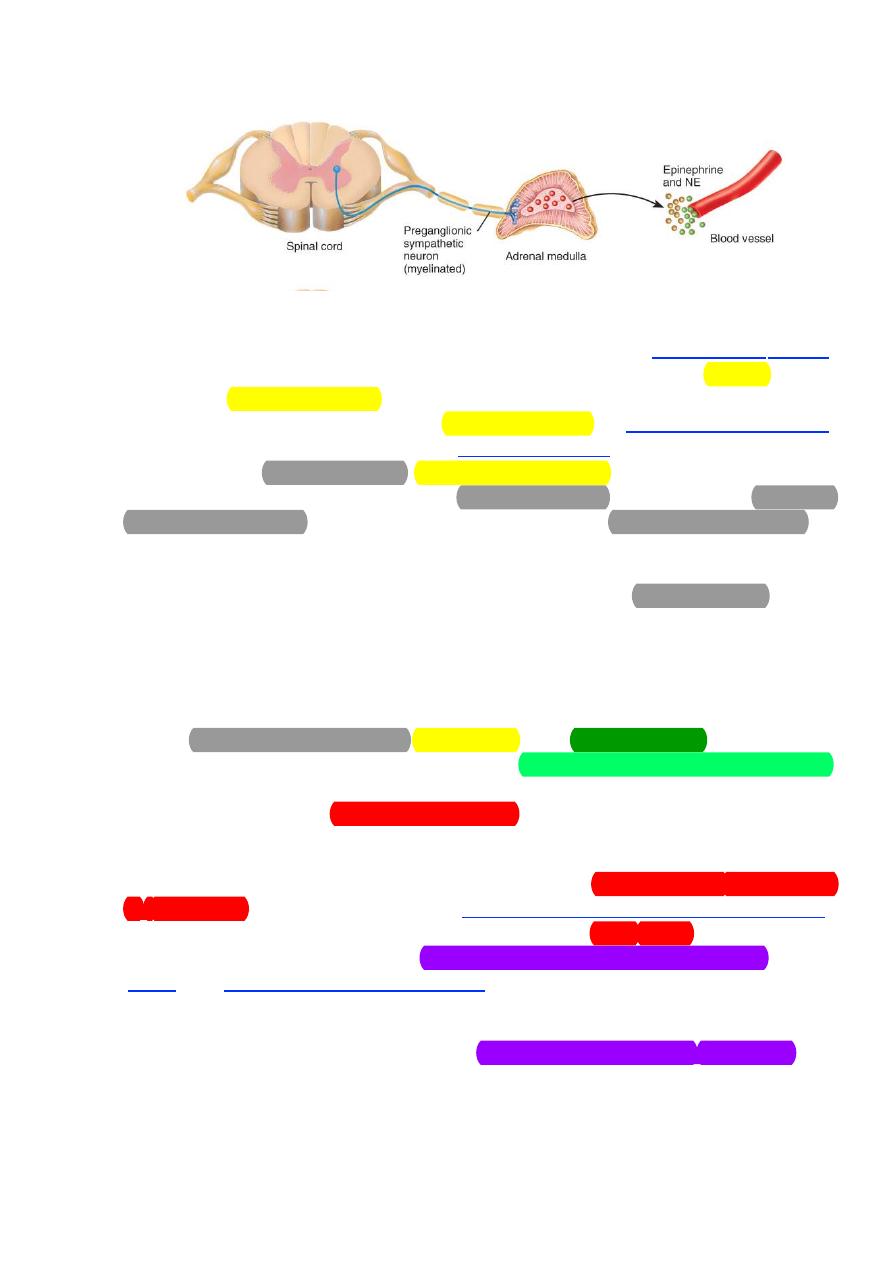
14
Figure 2: The direct release of NE to the blood.
●
The duration of activity of the catecholamines is significantly longer
than that of neuronally released norepinephrine. Therefore, the effects on the
tissues are more prolonged. This difference has to do with the mechanism of
inactivation of these substances. Norepinephrine is immediately removed
from the neuroeffector synapse by way of reuptake into the postganglionic
neuron. This rapid removal limits the duration of the effect of this
neurotransmitter. In contrast, there are no enzymes in the blood to degrade
the catecholamines. Instead, the catecholamines are inactivated by COMT in
the liver. As one might expect, the hepatic clearance of these hormones from
the blood would require several passes through the circulation. Therefore, the
catecholamines are available to cause their effects for a comparatively longer
period of time (up to 1-2 minutes as opposed to milliseconds).
●
Because they travel in the blood, organs and tissues throughout the
body are exposed to the catecholamines. Therefore, they are capable of
stimulating tissues that are not directly innervated by sympathetic nerve
fibers: airways smooth muscle, hepatocytes, and adipose tissue, in particular.
As a result, the catecholamines have a much wider breadth of activity
compared to norepinephrine released from sympathetic nerves. It also causes
a great increase in the basal metabolic rate (BMR).
●
The third important feature that distinguishes the catecholamines
from neuronally released norepinephrine involves epinephrine's affinity for
β
2
receptors. Norepinephrine has a very limited affinity for these receptors.
Therefore, circulating epinephrine causes effects that differ from those of
direct sympathetic innervations including a greater stimulatory effect on the
heart and relaxation of smooth muscle (vascular, bronchial, gastrointestinal,
and genitourinary).
Epinephrine and norepinephrine have equal affinity for β
1
receptors, the
predominant adrenergic receptor on the heart. However, the human heart
also contains a small percentage of β
2
receptors which, like β
1
receptors are

15
excitatory. Therefore, epinephrine is capable of stimulating a greater number
of receptors and of causing a greater stimulatory effect on the myocardium.
Beta two adrenergic receptors are also found on smooth muscle in
several organ systems. These receptors tend to be inhibitory and cause
relaxation of the smooth muscle. Vascular smooth muscle in skeletal muscle
contains both α
1
and β
2
receptors. Norepinephrine, which stimulates only the
excitatory α
1
receptors, causes strong vasoconstriction. However,
epinephrine, which stimulates both types of receptors, causes only weak
vasoconstriction. The vasodilatation resulting from β
2
receptor stimulation
opposes and, therefore, weakens the vasoconstriction resulting from α
1
receptor stimulation. Given that skeletal muscle may account for 40% of an
adult's body weight, the potential difference in vasoconstriction, blood
pressure, and the distribution of blood flow could be quite significant.
Another noteworthy example of the relaxation of smooth muscle by way
of β
2
receptor stimulation involves the airways. Bronchodilation, or the
opening of the airways, facilitates airflow in the lungs. Any direct sympathetic
innervation to the lungs is irrelevant in this respect, as only circulating
epinephrine is capable of stimulating these receptors on airway smooth
muscle.
Synthesis of NE:
Tyrosine
═
(hydroxylation)
═►
DOPA
═
(decarboxylation)
══►
Dopamine transport in vesicles ═ (hydroxylation) ═►nor epinephrine ═
(methylation) ═► Epinephrine
When NE or E come in contact with receptors on cell membrane of the
effector cells → receptor transmitter complex →activated enzyme adenyl
cyclase → cyclic AMP → Phosphorylation of Voltage-dependent Ca
++
channels
→ increase intracellular Ca
++
.
Sympathetic and parasympathetic tones:
The ANS is continuously active. The basal rate of activity is called
"Tone". The tone can be increased or decreased.
16
Interactions between the Sympathetic and
Parasympathetic Divisions:
Most visceral organs receive dual innervation—they are
innervated by both sympathetic and parasympathetic fibers. In this
condition, the effects of the two divisions of the autonomic system
may be antagonistic, complementary, or cooperative
Antagonistic Effects
The effects of sympathetic and parasympathetic innervation of
the pacemaker region of the heart is the best example of the
antagonism of these two systems. In this case, sympathetic and
parasympathetic fibers innervate the same cells. Adrenergic
stimulation from sympathetic fibers increases the heart rate,
whereas the release of acetylcholine from parasympathetic fibers
decreases the heart rate. A reverse of this antagonism is seen in the
digestive tract, where sympathetic nerves inhibit and
parasympathetic nerves stimulate intestinal movements and
secretions.
Complementary and Cooperative Effects
The effects of sympathetic and parasympathetic nerves are
generally antagonistic; in a few cases, however, they can be
complementary or cooperative. The effects are complementary
when sympathetic and parasympathetic stimulation produce similar
effects. The effects are cooperative, or synergistic, when sympathetic
and parasympathetic stimulation produce different effects that work
together to promote a single action.
The effects of sympathetic and parasympathetic stimulation on
salivary gland secretion are complementary. The secretion of watery

17
saliva is stimulated by parasympathetic nerves, which also stimulate
the secretion of other exocrine glands in the digestive tract.
Sympathetic nerves stimulate the constriction of blood vessels
throughout the digestive tract. The resultant decrease in blood flow
to the salivary glands causes the production of thicker, more viscous
saliva.
The effects of sympathetic and parasympathetic stimulation on
the reproductive and urinary systems are cooperative.
Erection of the penis, for example, is due to vasodilation
resulting from parasympathetic nerve stimulation; ejaculation is due
to stimulation through sympathetic nerves. The two divisions of the
autonomic system thus cooperate to enable sexual function in the
male.
Organs without Dual Innervation
Although most organs are innervated by both sympathetic and
parasympathetic nerves, some—including the adrenal medulla,
arrector pili muscles, sweat glands, and most blood vessels—receive
only sympathetic innervation. In these cases, regulation is achieved
by increases or decreases in the tone (firing rate) of the sympathetic
fibers. Constriction of cutaneous blood vessels, for example, is
produced by increased sympathetic activity that stimulates alpha-
adrenergic receptors, and vasodilation results from decreased
sympathetic nerve stimulation.
The sweat glands in the trunk secrete a watery fluid in
response to cholinergic sympathetic stimulation. Evaporation of this
dilute sweat helps to cool the body. At the end of exercise,
sympathetic stimulation is reduced, causing cutaneous blood vessels
to dilate. This increases blood flow to the skin, which helps to
eliminate metabolic heat.
Regulation of Autonomic Nervous System Activity
The efferent nervous activity of the ANS is largely regulated by
autonomic reflexes. In many of these reflexes, sensory information is
transmitted to homeostatic control centers, in particular, those located in the
hypothalamus and brainstem. Much of the sensory input from the thoracic
and abdominal viscera is transmitted to the brainstem by afferent fibers of
cranial nerve X, the vagus nerve. Other cranial nerves also contribute sensory
input to the hypothalamus and the brainstem. This input is integrated and a
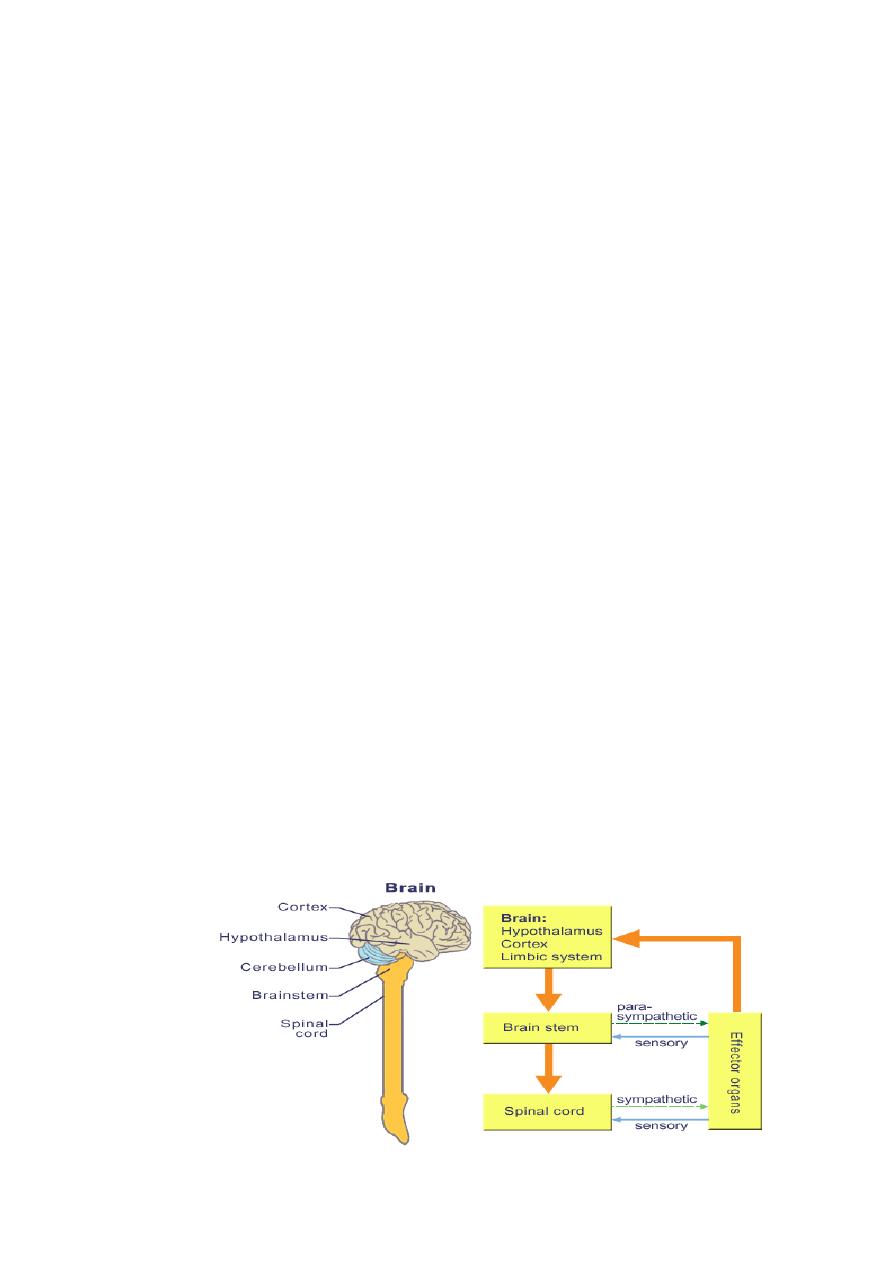
18
response is carried out by the transmission of nerve signals that modify the
activity of preganglionic autonomic neurons. Many important variables in the
body are monitored and regulated in the hypothalamus and the brainstem
including heart rate, blood pressure, gastrointestinal peristalsis and glandular
secretion, body temperature, hunger, thirst, plasma volume, and plasma
osmolarity.
An example of this type of autonomic reflex is the baroreceptor reflex.
Baroreceptors located in some of the major systemic arteries are sensory
receptors that monitor blood pressure. If blood pressure decreases, the
number of sensory impulses transmitted from the baroreceptors to the
vasomotor center in the brainstem also decreases. As a result of this change in
baroreceptor stimulation and sensory input to the brainstem, ANS activity to
the heart and blood vessels is adjusted to increase heart rate and vascular
resistance so that blood pressure increases to its normal value.
These neural control centers in the hypothalamus and the brainstem
may also be influenced by higher brain areas. Specifically, the cerebral cortex
and the limbic system influence ANS activities associated with emotional
responses by way of hypothalamic-brainstem pathways. For example,
blushing during an embarrassing moment, a response most likely originating
in the frontal association cortex, involves vasodilation of blood vessels to the
face. Other emotional responses influenced by these higher brain areas
include fainting, breaking out in a cold sweat, and a racing heart rate.
Some autonomic reflexes may be processed at the level of the spinal
cord. These include the micturition reflex (urination) and the defecation
reflex. Although these reflexes are subject to influence from higher nervous
centers, they may occur without input from the brain.
End