Liver, Gallbladder, and Pancreas
OBJECTIVESThese lecture notes should help the student to:
- Describe the classic liver lobule, the portal lobule, and the hepatic acinus. Understand the functions that gave rise to these overlapping views of liver organization.
- Relate the hepatocyte’s complex ultrastructure to its many functions.
- Describe the liver’s double blood supply.
- Describe the hepatic sinusoids.
- Describe the location, contents, and function of the space of Disse.
- Describe the biliary tract.
- Describe the structure, function, and location of the gallbladder.
- Relate the ultrastructure of the pancreatic acinar cell to its function.
- Describe the structure and function of the islets of Langerhans.
- Distinguish between the structure of the pancreas and parotid gland.
Histology
Prof.Dr.Faraid Lec.5 LIVERThe liver is located in the peritoneal cavity below the diaphragm. The liver is the largest gland in the body and is the second largest organ (the largest is the skin), weighing about 1.5 Kg in the adult. The liver produces bile and plays a major role in lipid, carbohydrate, and protein metabolism. The liver inactivates and metabolizes many toxic substances and drugs. It also participates in iron metabolism and the synthesis of blood proteins and the factors necessary for blood coagulation. Elimination of toxic substances occurs in the bile, an exocrine secretion of the liver that is important for lipid digestion.
The liver is covered by a thin connective tissue capsule (Glisson's capsule) that becomes thicker at the hilum (porta hepatis) where the portal vein and the hepatic artery enter the organ and where the right and the left hepatic ducts and lymphatics exit. Trabeculae arise from the capsule subdivide the liver into lobes and lobules. In man the trabeculae are very incomplete.
The structural units of the liver are called liver lobules. In certain animals (eg. pigs) these lobules are separated from each other and sharply delimited by a layer of connective tissue. This is not the case in humans, making it difficult to establish the exact limits between different lobules.
Liver Lobules:
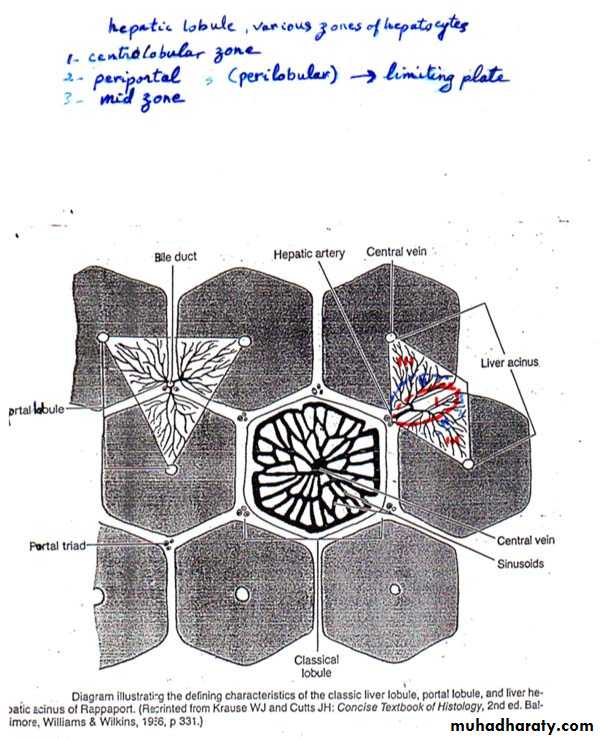
1. Classic Liver Lobule: (this is based on the direction of the blood flow)
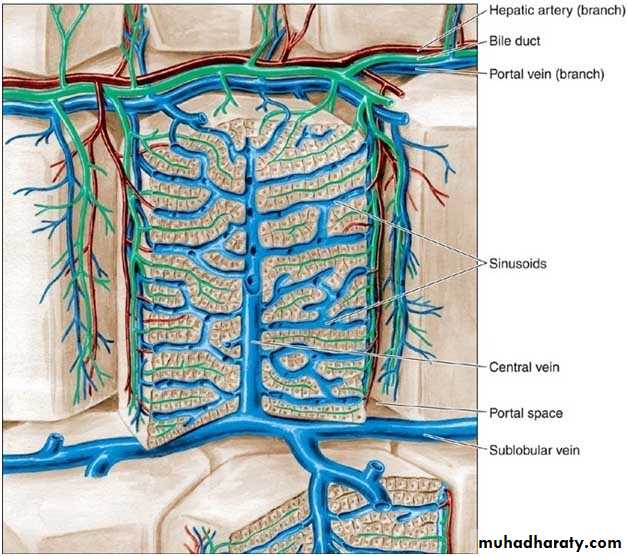
It is a hexagonal mass of tissue primarily composed of plates of hepatocytes which radiate from the region of the central vein (present in the center) toward the periphery. The plates are separated by hepatic sinusoids. At the corners of the lobule, there is the portal canals [portal areas, portal tracts, portal spaces or portal triad].Each portal canal contains: a portal venule (a branch of portal vein), a hepatic arteriole (a branch of hepatic artery), bile duct and lymphatic vessels. These structures are surrounded by loose connective tissue.
The arterial and venous blood flows centripetally in each lobule (from the portal areas to drain in the central vein), whereas the bile flows centrifugally towards the portal areas.
In the lobule, various zones of hepatocytes can be identified. These are the centrolobular, periportal, and mid zones.
The centrolobular zone: is composed of hepatocytes surrounding the central vein, and is the zone most distant from the oxygenated arterial blood supply.
The periportal (perilobular zone): at the periphery of the lobule, is closely related to the portal tracts. The outermost layer of periportal hepatocytes immediately adjacent to the portal tract is called the limiting plate; it is the first group of hepatocytes to be damaged in liver disorders that primarily involve the portal tracts.
The mid zone: is the zone of hepatocytes between the centrolobular and periportal zones.
2. Portal Lobule: (this is based mainly on the direction of bile flow)
It is a triangular region with a portal triad at its center and a central vein at each of its three corners. It contains portions of three adjacent classic liver lobules all of which drain bile into one portal canal.
3. Hepatic Acinus (of Rappaport): (it is based on changes in oxygen, nutrient, and toxin content as blood flowing through the sinusoids is acted on by hepatocytes)
It is that region which is supplied by a terminal branch of the portal vein and hepatic artery and drained by a terminal branch of the bile duct. It is a diamond-shaped region, contains portions (triangular sections) of two adjacent classic liver lobules (whose apices are the central veins).
In relation to their proximity to the distributing vessels, cells in the hepatic acinus can be subdivided into three zones. Cells in zone I would be those closest to the distributing vessels, whereas zone III is closer to the central vein. Zone II is intermediate between zone I and III. Blood in zone I sinusoids has higher oxygen, nutrient, and toxin concentrations than in the other zones. As the blood flows toward the central vein, these substances are gradually removed by hepatocytes. Zone I hepatocytes thus have a higher metabolic rate and larger glycogen and lipid stores. They are also more susceptible to damage by blood-borne toxins, and their energy stores are the first to be depleted during fasting. This explains regional histopathologic differences in patients with liver damage.
Hepatocytes:
They are the main cells of the lobules. They are epithelial cells (derived from embryonic endoderm).
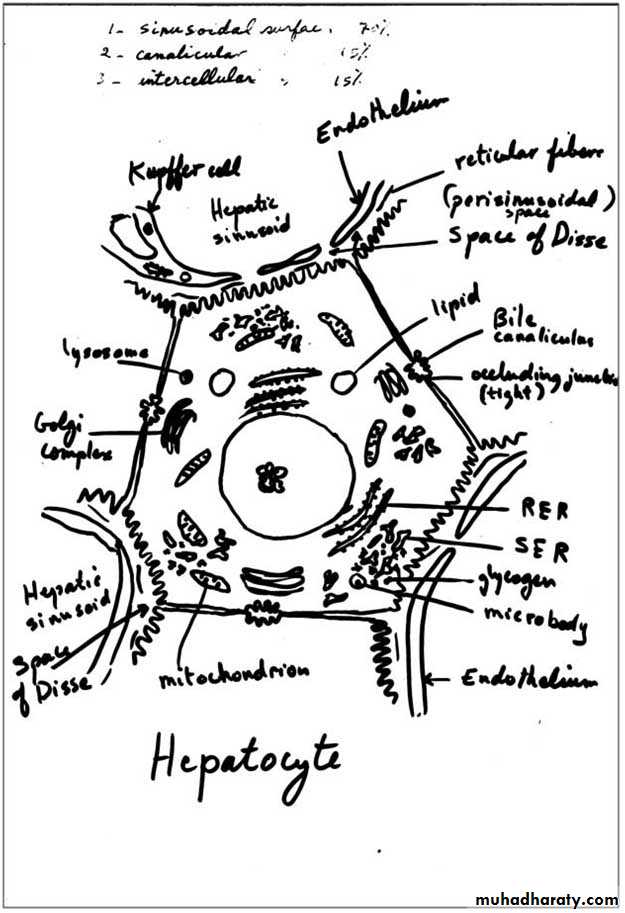
They are arranged in plates that anastomose freely and separated from each other by spaces filled by hepatic sinusoids and are disposed radially around the central vein. Hepatocytes are large polyherdral cells (20-30 µm in diameter). Hepatocytes have three important surfaces:
Sinusoidal surfaces: are separated from the sinusoidal vessel by the space of Disse. They account for approximately 70% of the total hepatocyte surface. They are covered by short microvilli, which protrude into the space of Disse. The sinusoidal surface is the site where material is transferred between the sinusoids and the hepatocyte.
Canalicular surfaces: are the surfaces across which bile drains from the hepatocytes into the canaliculi. They account for approximately 15% of the hepatocyte surface.
The intercellular surfaces: are the surfaces between adjacent hepatocytes that are not in contact with sinusoids or canaliculi. They account for about 15% of the hepatocyte surface.
Hepatocytes usually contain one round, central nucleus; some are binucleated (25%). Occasionally nuclei are polyploid (more than one set of chromosomes). The cytoplasm of the hepatocytes is typically fairly acidophilic (many mitochondria and some SER), with areas of basophilia (rough endoplasmic reticulum). The hepatocytes are unusual in that they possess abundant RER and SER in the same cells. The RER is associated with protein synthesis such as: albumin, fibrinogen, globulin, prothrombin. The SER is associated with steroid metabolism and also is responsible for the processes of oxidation, methylation, and conjugation required for inactivation or detoxification of various substances before their excretion from the body.
One of the main processes occurring in the SER is the conjugation of water-insoluble toxic bilirubin by glucuronyl-transferase to form a water-soluble nontoxic bilirubin glucuronide. This conjugate is excreted by hepatocytes into the bile. When bilirubin or bilirubin glucuronide is not excreted, various diseases characterized by jaundice can result.
Each hepatocyte has approximately 2000 mitochondria. Golgi complexes are also numerous up to 50 per cell. The functions of this organelle include the formation of lysosomes and the secretion of plasma proteins (eg. albumin, proteins of the complement system), glycoproteins (eg. transferrin), and lipoproteins (eg. very low-density lipoproteins). Hepatocytes possess lysosomes which are important in the turnover and degradation of interacellular organelles. Peroxisomes are abundant. Lipid droplet and glycogen are found in varying amounts depending upon the functional state of the cells. The glycogen appears in the electron microscope as coarse, electron dense granules that frequently collect close to the SER.
The hepatocyte is probably the most versatile cell in the body. It is a cell with both endocrine and exocrine functions. Endocrine secretion involves the production and release of several plasma proteins. Unlike other protein synthesizing and secreting cells of the body, the hepatocytes are unusual in that they lack protein storage granules. Exocrine secretion involves the production and release of bile.
Hepatocytes located at different distances from the portal triads show differences in structural, histochemical, and biochemical characteristics.
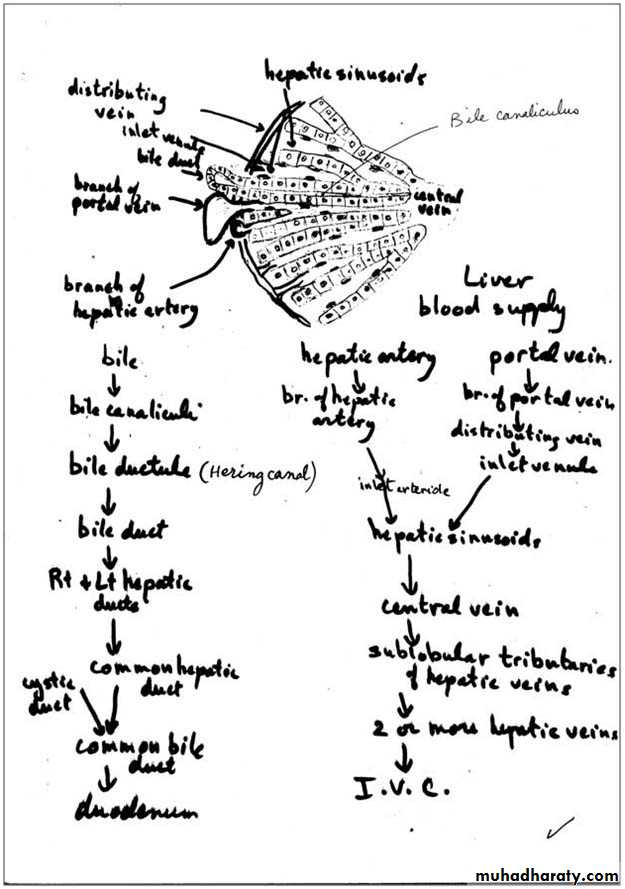
Bile canaliculi: are the first portions of the bile duct system. They are tubular spaces 1-2 µm in diameter, limited only by the plasma membranes of 2 adjacent hepatocytes and have a small number of microvilli in their interiors. The cell membranes near these canaliculi are sealed together by tight junctions. The canaliculi drain outwards in the direction of the portal canal. The bile flow therefore progresses in a direction opposite to that of the blood, i.e., from the center of the lobule to its periphery. At the periphery, bile enters the bile ductules (or Hering's canals), lined with cuboidal epithelium. After a short distance, the ductules cross the limiting hepatocytes of the lobule and end in the bile ducts in the portal spaces. Bile ducts are lined by cuboidal or columnar epithelium and have a distinct c.t. sheath. They gradually enlarge and fuse, forming right and left hepatic ducts, which leave the liver, unite to form a common hepatic duct. The hepatic duct after receiving the cystic duct from the gallbladder continues to the duodenum as the common bile duct. The hepatic, cystic, and common bile ducts are lined with simple columnar epithelium.
Hepatic sinusoids: are vessels that arise at the periphery of a lobule and run between adjacent plates of hepatocytes. They receive blood from the vessels in the portal areas and deliver it to the central vein. Sinusoids are larger than capillaries, more irregular in shape and their lining cells are directly related to the epithelial cells with no intervening c.t. The lining of the sinusoids consists of a discontinuous layer of fenestrated endothelium also contain Kupffer cells in their endothelial lining. They lack basal lamina.
Kupffer cells: are mononuclear phagocytic cells (fixed macrophages) derived from blood monocytes and located in the lining of hepatic sinusoids. They are large cells with several processes, and exhibit an irregular or stellate outline. Their main functions are to metabolize aged erythrocytes, digest hemoglobin, secrete proteins related to immunologic processes, and destroy bacteria that enter the portal blood through the large intestine.
Space of Disse: is the perisinusoidal space located between hepatocytes and the endothelium of hepatic sinusoids. It contains microvilli of the hepatocytes, plasma, reticular fibers, and fat-storing cells (Ito cells). Blood plasma enters this space through openings between the endothelial cells that are too small for blood cells to pass. Blood-borne substances thus directly contact the microvilli of hepatocytes. These cells absorb nutrients, oxygen, and toxins from, and release endocrine secretions into, these spaces.
It functions in the exchange of material between the bloodstream and hepatocytes, which do not directly contact the blood stream.
Fat-storing (Ito) cells: are stellate cells lie in the space of Disse and have the ability to accumulate lipid droplets. They are the main source of vitamin A storage in the body.
Blood supply of the liver:
The liver is unusual in that it receives blood from two sources:70- 80% of the blood derives from the portal vein, which carries oxygen-poor, nutrient-rich blood from the abdominal viscera, and 20-30% derives from the hepatic artery, which supplies oxygen-rich blood.
Portal vein system:
The portal vein branches repeatedly and sends small portal venules to the portal spaces. The portal venules branch into the distributing veins that run around the periphery of the lobule. From the distributing veins, small inlet venules empty into the sinusoids. The sinusoids run radially, converging in the center of the lobule to form the central, or centrolobular, vein. The central vein has thin walls consisting only of endothelial cells supported by a sparse population of collagen fibers. As the central vein progresses along the lobule, it receives more and more sinusoids and gradually increases in diameter. At its end, it leaves the lobule at its base by merging with the larger sublobular vein. The sublobular veins gradually converge and fuse, forming the two or more large hepatic veins that empty into the inferior vena cava.
The portal system conveys blood from the pancreas and spleen and blood containing nutrients absorbed in the intestines. The portal vein is formed by the junction of mesenteric and splenic veins.
Arterial system:
The hepatic artery branches repeatedly and forms the interlobular arteries (hepatic arterioles). Some of these arteries irrigate the structures of the portal canal, and others form arterioles (inlet arterioles) that end directly in the sinusoids, thus providing a mixture of arterial and portal venous blood in the sinusoids. The hepatic artery is a branch of the celiac artery of the abdominal aorta.
Blood flows from the periphery to the center of the liver lobule. Consequently, oxygen and metabolites, as well as all other toxic or nontoxic substances absorbed in the intestines, reach the peripheral cells first and then reach the central cells of the lobule. This direction of blood flow explains why the behavior of perilobular cells differs from that of the centrolobular cells. This is particularly evident in pathologic specimens, where changes are seen in either the central cells or the peripheral cells of the lobule.
Liver Regeneration:
Despite its low rate of cell renewal, the liver has an extraordinary capacity for regeneration. The process of regeneration is probably controlled by circulating substances called chalones, which inhibit the mitotic division of certain cell types. When a tissue is injured or partially removed, the number of chalones it produces decreases; consequently, a burst of mitotic activity occurs in this tissue. As regeneration proceeds, the quantity of chalones produced increases and mitotic activity decreases. This process is self-regulating. The regenerated liver tissue is usually similar to the removed tissue. If there is continuous or repeated damage to hepatocytes over a long period, the multiplication of liver cells is followed by a pronounced increase in the amount of connective tissue. The excess of c.t. results in disorganization of the liver structure, a condition known as cirrhosis, is a progressive and irreversible process, causes liver failure, and is usually fatal.
Cirrhosis is a consequence of any sustained progressive injury to hepatocytes produced by several agents, such as ethanol, drugs or other chemicals, hepatitis virus, and autoimmune liver disease.
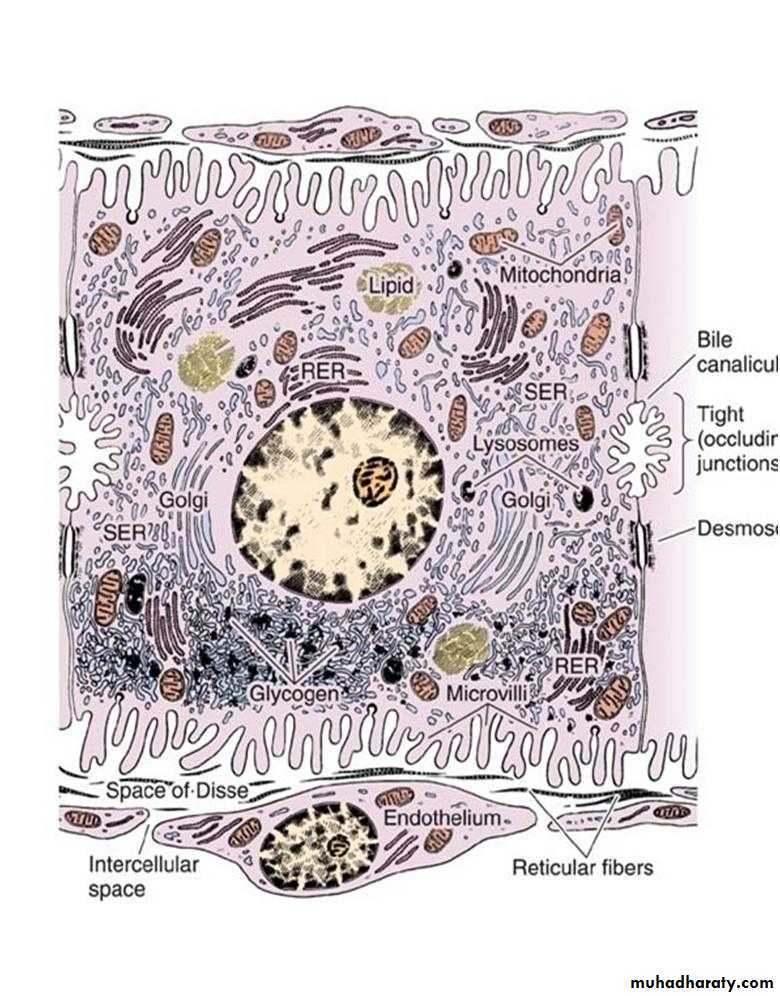
Ultrastructure of a hepatocyte. RER, rough endoplasmic reticulum; SER, smooth endoplasmic reticulum. x10,000
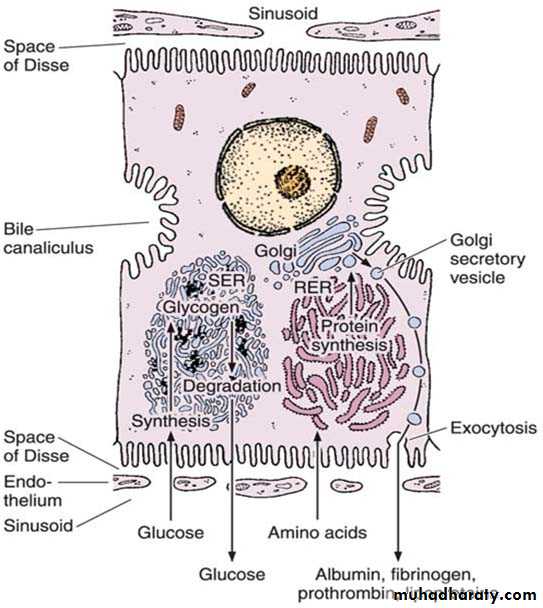
Protein synthesis and carbohydrate storage in the liver. Carbohydrate is stored as glycogen, usually associated with the smooth endoplasmic reticulum (SER). When glucose is needed, glycogen is degraded. In several diseases, glycogen degradation is depressed, resulting in abnormal intracellular accumulations of glycogen. Proteins produced by hepatocytes are synthesized in the rough endoplasmic reticulum (RER), which explains why hepatocyte lesions or starvation lead to a decrease in the amounts of albumin, fibrinogen, and prothrombin in a patient’s blood. The impairment of protein synthesis leads to several complications, since most of these proteins are carriers, important for the blood’s osmotic pressure and for coagulation.
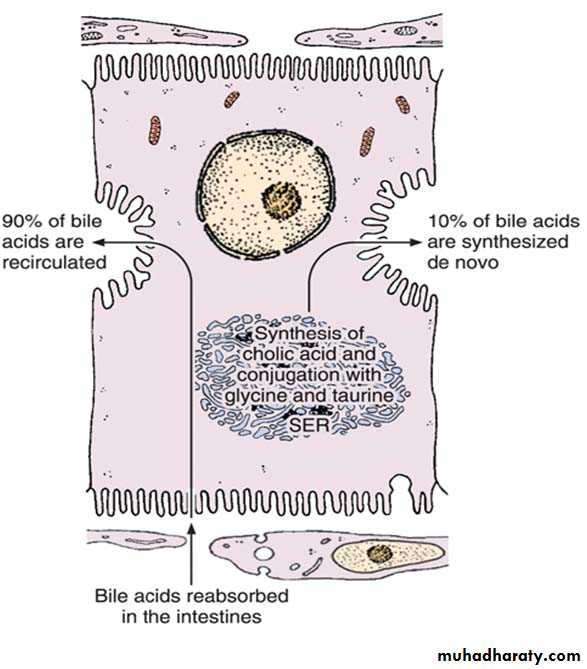
Mechanism of secretion of bile acids. About 90% of bile acids are derived from the intestinal epithelium and transported to the liver. The remaining 10% are synthesized in the liver by the conjugation of cholic acid with the amino acids glycine and taurine. This process occurs in the smooth endoplasmic reticulum (SER).
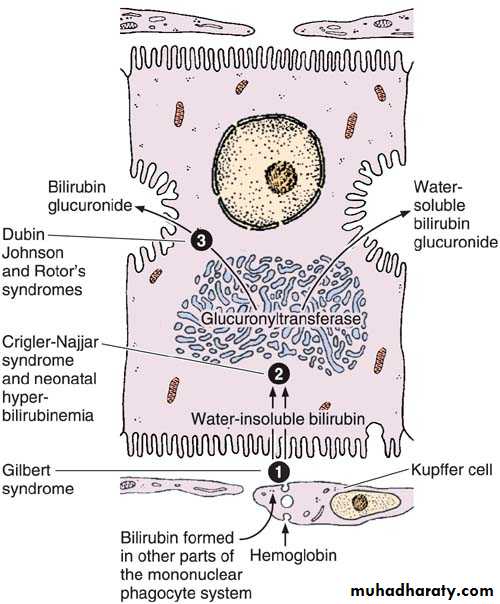
The secretion of bilirubin. The water-insoluble form of bilirubin is derived from the metabolism of hemoglobin in macrophages. Glucuronyltransferase activity in the hepatocytes causes bilirubin to be conjugated with glucuronide in the smooth endoplasmic reticulum, forming a water-soluble compound. When bile secretion is blocked, the yellow bilirubin or bilirubin glucuronide is not excreted; it accumulates in the blood, and jaundice results. Several defective processes in the hepatocytes can cause diseases that produce jaundice: a defect in the capacity of the cell to trap and absorb bilirubin (1), the inability of the cell to conjugate bilirubin because of a deficiency in glucuronyltransferase (2), or problems in the transfer and excretion of bilirubin glucuronide into the bile canaliculi (3). One of the most frequent causes of jaundice, however–unrelated to hepatocyte activity–is the obstruction of bile flow as a result of gallstones or tumors of the pancreas.
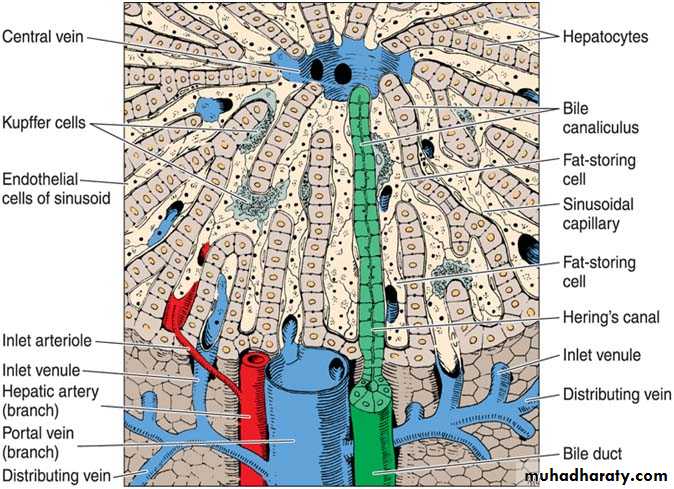
Three-dimensional aspect of the normal liver. In the upper center is the central vein; in the lower center is the portal vein. Note the bile canaliculus, liver plates, Hering’s canal, Kupffer cells, sinusoid, fat-storing cell, and sinusoid endothelial cells. (Courtesy of M Muto.)
Schematic drawing of the structure of the liver. The liver lobule in the center is surrounded by the portal space (dilated here for clarity). Arteries, veins, and bile ducts occupy the portal spaces. Nerves, connective tissue, and lymphatic vessels are also present but are (again, for clarity) not shown in this illustration. In the lobule, note the radial disposition of the plates formed by hepatocytes; the sinusoidal capillaries separate the plates. The bile canaliculi can be seen between the hepatocytes. The sublobular (intercalated) veins drain blood from the lobules. (Redrawn and reproduced, with permission, from Bourne G: An Introduction to Functional Histology. Churchill, 1953.
Histology
Prof.Dr.Faraid Lec.6Gallbladder:
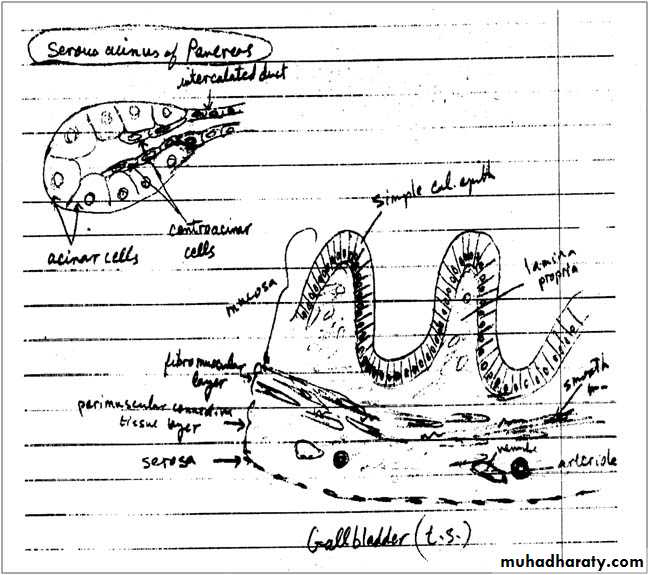
It is a hollow, pear-shaped organ attached to the lower surface of the liver. It can store 30-50 ml of bile. The gallbladder absorbs water from the bile and stores the bile in a concentrated form. The wall of the gallbladder consists of the following layers:
Mucosa: is composed of a simple columnar epithelium and a richly vascularized lamina propria. The mucous membrane is thrown into folds that are particularly evident in the empty bladder. There is no gland in the gallbladder except in the neck region.
Fibromuscular layer: is composed of a thin layer of smooth muscle fibers interspersed within the layers of loose c.t. that are rich in elastic fibers. The muscle contracts and empties the gallbladder in response to cholecystokinin released by entero endocrine cells (I-cells) in the intestinal mucosa when dietary fat enters the small intestine.
Perimuscular connective tissue layer: is a wide layer of connective tissue, which contains blood vessels, lymphatic and nerves.
A thick layer of c.t. binds the superior surface of the gallbladder to the liver.
Serosa: covers its free surface.
Pancreas:
The pancreas is a mixed exocrine-endocrine gland that produces digestive enzymes and hormones. The enzymes are stored and released by cells of the exocrine portion. The hormones are synthesized in clusters of endocrine epithelial cells known as islets of Langerhans. The pancreas is a retroperitoneal gland. It has a head, body, and tail. The head is lodged in the concavity of the C-shaped duodenum and its narrower boy and tail extend to the hilus of the spleen.
A thin capsule of c.t. covers the pancreas and sends septa into it, separating the pancreatic lobules. These septa convey blood and lymphatic vessels into and out of the parenchyma and house the interlobular ducts.
The exocrine portion of the pancreas is a compound acinar serous gland, similar in structure to the parotid gland. In histologic sections, a distinction between the two glands can be made based on the absence of striated ducts and the presence of the islets of Langerhans in the pancreas. Another characteristic is that in the pancreas the initial portions of intercalated ducts penetrate the lumens of the acini. Nuclei, surrounded by a pale cytoplasm, belong to centroacinar cells that constitute the intra-acinar portion of the intercalated duct. Centroacinar cells are found only in the pancreatic acini. Intercalated ducts are tributaries of larger interlobular ducts. There are no striated ducts in the pancreatic ducts system.
The exocrine pancreatic acinus is composed of several serous cells surrounding a lumen. These cells are highly polarized, with a spherical nucleus, and are typical protein-secreting cells. The number of zymogen (secretory) granules present in each cell varies according to the digestive phase and attains its maximum in animals that have fasted. Myoepithelial cells do not surround the acini in the pancreas.
The human exocrine pancreas secretes, besides water and ions, the following enzymes and proenzymes: trypsinogens, chymotrypsinogen, carboxy peptidases, ribonuclease, deoxyribonuclease, amylase, lipase, phospholipase and elastase. The majority of the enzymes are stored as proenzymes in the secretory granules of acinar cells, being activated in the lumen of the small intestine after secretion. This is very important for the protection of the pancreas. In acute pancreatitis, the proenzymes may be activated and digest the whole pancreas, leading to very serious complications.
Pancreatic secretion is controlled mainly through two hormones secretin and cholecystokinin that are produced by enteroendocrine cells of the duodenal mucosa. Stimulation of the vagus nerve (parasympathetic stimulation) will also produce pancreatic secretion.
Secretin promotes secretion of an abundant fluid, poor in enzyme activity and rich in bicarbonate.
Cholecystokinin promotes secretion of a less abundant but enzyme-rich fluid.
Islets of Langerhans (endocrine pancreas):
They appear as rounded clusters of cells embedded within exocrine pancreatic tissue. Most islets are 100-200 µm in diameter and contain several hundred cells, small islets also are found. There may be more than one million islets in the human pancreas, with a slight tendency for islets to be more abundant in the tail region.
In sections, each islet consists of lightly stained polygonal or rounded cells, arranged in cords separated by a network of blood capillaries. A fine capsule of reticular fibers surrounds each islet.
Using immunocytochemical methods four types of cells A, B, D, and F have been located in the islets. The ultrastructure of these cells resembles that of cells synthesizing polypeptides.
A or α-cells ≈ 20% produce the hormone glucagon whose effects are opposite those of insulin. They are located mostly at periphery of islets. By using special stain, the granules in their cytoplasm stain acidophilic.
B or β-cells are the most numerous ≈ 70%. Produce insulin and tend to be concentrated in the center of the islet their granules stain blue.
D or delta cells 5% produce somatostatin.
F or PP cells 1-2% secrete pancreatic polypeptide.
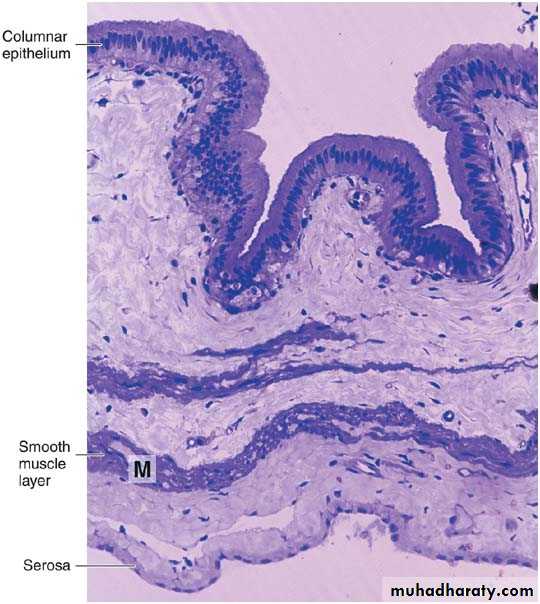
Photomicrograph of a section of gallbladder. Note the lining of columnar epithelium and the smooth muscle layer (M). PT stain. Low magnification.
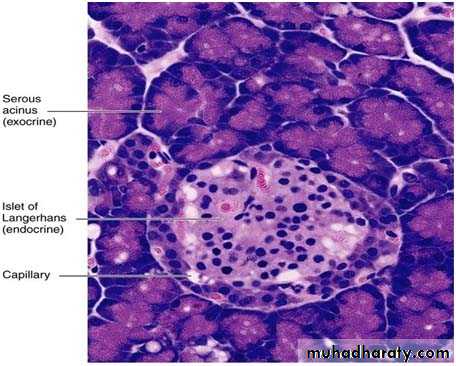
Photomicrograph of a pancreas showing the exocrine portion (acini) and the endocrine portion (islet of Langerhans). The acini contain secretory cells with basophilic cytoplasm. Different types of endocrine cells are seen in the islet. PT stain. Medium magnification.
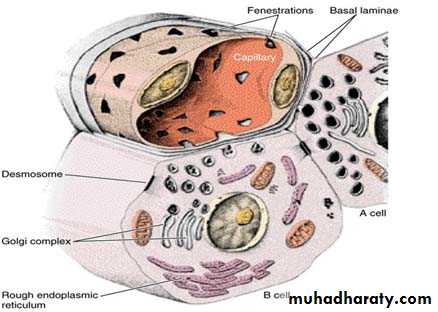
Drawing of the A and B cells; showing their main ultrastructural features. The B cell’s granules are irregular, whereas the A cell’s granules are round and uniform.