The Female Reproductive System
PART IILecture 4
29-2-2016Functions of the Ovarian Hormones-Estradiol and Progesterone
The two types of ovarian sex hormones are the estrogens and the progestins. By far the most important of the estrogens is the hormone estradiol, and by far the most important progestin is progesterone. The estrogens mainly promote proliferation and growth of specific cells in the body that are responsible for the development of most secondary sexual characteristics of the female. The progestins function mainly to prepare the uterus for pregnancy and the breasts for lactation.Chemistry of the Sex Hormones
EstrogensIn the normal nonpregnant female, estrogens are secreted in significant quantities only by the ovaries, although minute amounts are also secreted by the adrenal cortices. During pregnancy, tremendous quantities of estrogens are also secreted by the placenta, as discussed in Chapter 82.
Only three estrogens are present in significant quantities in the plasma of the human female: β-estradiol, estrone, and estriol, the formulas for which are shown in Figure 81-6. The principal estrogen secreted by the ovaries is β-estradiol. Small amounts of estrone are also secreted, but most of this is formed in the peripheral tissues from androgens secreted by the adrenal cortices and by ovarian thecal cells. Estriol is a weak estrogen; it is an oxidative product derived from both estradiol and estrone, with the conversion occurring mainly in the liver.
The estrogenic potency of β-estradiol is 12 times that of estrone and 80 times that of estriol. Considering these relative potencies, one can see that the total estrogenic effect of β-estradiol is usually many times that of the other two together. For this reason, β-estradiol is considered the major estrogen, although the estrogenic effects of estrone are not negligible.
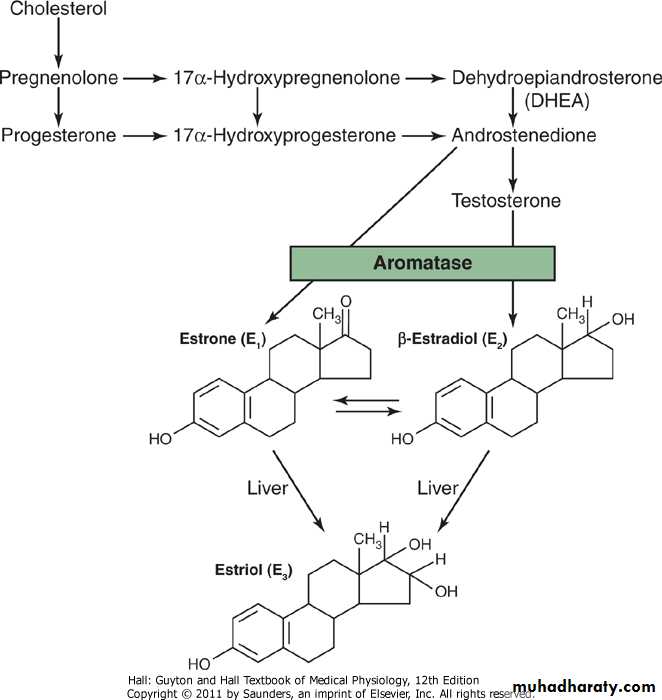
Figure 81-6 Synthesis of the principal female hormones. The chemical structures of the precursor hormones, including progesterone, are shown in Figure 77-2.
Progestins
By far the most important of the progestins is progesterone. However, small amounts of another progestin, 17-α-hydroxyprogesterone, are secreted along with progesterone and have essentially the same effects. Yet for practical purposes, it is usually reasonable to consider progesterone the only important progestin.In the normal nonpregnant female, progesterone is secreted in significant amounts only during the latter half of each ovarian cycle, when it is secreted by the corpus luteum.
As we shall see in Chapter 82, large amounts of progesterone are also secreted by the placenta during pregnancy, especially after the fourth month of gestation.
Synthesis of the Estrogens and Progestins
Note from the chemical formulas of the estrogens and progesterone in Figure 81-6 that they are all steroids. They are synthesized in the ovaries mainly from cholesterol derived from the blood but also to a slight extent from acetyl coenzyme A, multiple molecules of which can combine to form the appropriate steroid nucleus.Estrogens and Progesterone Are Transported in the Blood Bound to Plasma Proteins
Both estrogens and progesterone are transported in the blood bound mainly with plasma albumin and with specific estrogen- and progesterone-binding globulins. The binding between these hormones and the plasma proteins is loose enough that they are rapidly released to the tissues over a period of 30 minutes or so.Functions of the Liver in Estrogen Degradation
The liver conjugates the estrogens to form glucuronides and sulfates, and about one fifth of these conjugated products is excreted in the bile; most of the remainder is excreted in the urine. Also, the liver converts the potent estrogens estradiol and estrone into the almost totally impotent estrogen estriol. Therefore, diminished liver function actually increases the activity of estrogens in the body, sometimes causing hyperestrinism.
Fate of Progesterone
Within a few minutes after secretion, almost all the progesterone is degraded to other steroids that have no progestational effect. As with the estrogens, the liver is especially important for this metabolic degradation.The major end product of progesterone degradation is pregnanediol. About 10 percent of the original progesterone is excreted in the urine in this form. Therefore, one can estimate the rate of progesterone formation in the body from the rate of this excretion.
FUNCTIONS OF THE ESTROGENS
Their Effects on the Primary and Secondary Female Sex CharacteristicsA primary function of the estrogens is to cause cellular proliferation and growth of the tissues of the sex organs and other tissues related to reproduction.
Effect of Estrogens on the Uterus and External Female Sex Organs
During childhood, estrogens are secreted only in minute quantities, but at puberty, the quantity secreted in the female under the influence of the pituitary gonadotropic hormones increases 20-fold or more. At this time, the female sex organs change from those of a child to those of an adult. The ovaries, fallopian tubes, uterus, and vagina all increase several times in size. Also, the external genitalia enlarge, with deposition of fat in the mons pubis and labia majora and enlargement of the labia minora.In addition, estrogens change the vaginal epithelium from a cuboidal into a stratified type, which is considerably more resistant to trauma and infection than is the prepubertal cuboidal cell epithelium. Vaginal infections in children can often be cured by the administration of estrogens simply because of the resulting increased resistance of the vaginal epithelium.
During the first few years after puberty, the size of the uterus increases twofold to threefold, but more important than the increase in uterus size are the changes that take place in the uterine endometrium under the influence of estrogens. Estrogens cause marked proliferation of the endometrial stroma and greatly increased development of the endometrial glands, which will later aid in providing nutrition to the implanted ovum. These effects are discussed later in the chapter in connection with the endometrial cycle.
Effect of Estrogens on the Fallopian Tubes
The estrogens' effect on the mucosal lining of the fallopian tubes is similar to that on the uterine endometrium. They cause the glandular tissues of this lining to proliferate; especially important, they cause the number of ciliated epithelial cells that line the fallopian tubes to increase. Also, activity of the cilia is considerably enhanced. These cilia always beat toward the uterus, which helps propel the fertilized ovum in that direction.Effect of Estrogens on the Breasts
The primordial breasts of females and males are exactly alike. In fact, under the influence of appropriate hormones, the masculine breast during the first 2 decades of life can develop sufficiently to produce milk in the same manner as the female breast.Estrogens cause (1) development of the stromal tissues of the breasts, (2) growth of an extensive ductile system, and (3) deposition of fat in the breasts. The lobules and alveoli of the breast develop to a slight extent under the influence of estrogens alone, but it is progesterone and prolactin that cause the ultimate determinative growth and function of these structures.
In summary, the estrogens initiate growth of the breasts and of the milk-producing apparatus. They are also responsible for the characteristic growth and external appearance of the mature female breast. However, they do not complete the job of converting the breasts into milk-producing organs.
Effect of Estrogens on the Skeleton
Estrogens inhibit osteoclastic activity in the bones and therefore stimulate bone growth. As discussed in Chapter 79, at least part of this effect is due to stimulation of osteoprotegerin, also called osteoclastogenesis inhibitory factor, a cytokine that inhibits bone resorption.At puberty, when the female enters her reproductive years, her growth in height becomes rapid for several years. However, estrogens have another potent effect on skeletal growth: They cause uniting of the epiphyses with the shafts of the long bones. This effect of estrogen in the female is much stronger than the similar effect of testosterone in the male. As a result, growth of the female usually ceases several years earlier than growth of the male. A female eunuch who is devoid of estrogen production usually grows several inches taller than a normal mature female because her epiphyses do not unite at the normal time.
Osteoporosis of the Bones Caused by Estrogen Deficiency in Old Age
After menopause, almost no estrogens are secreted by the ovaries. This estrogen deficiency leads to (1) increased osteoclastic activity in the bones, (2) decreased bone matrix, and (3) decreased deposition of bone calcium and phosphate. In some women this effect is extremely severe, and the resulting condition is osteoporosis, described in Chapter 79. Because this can greatly weaken the bones and lead to bone fracture, especially fracture of the vertebrae, many postmenopausal women are treated prophylactically with estrogen replacement to prevent the osteoporotic effects.
Estrogens Slightly Increase Protein Deposition
Estrogens cause a slight increase in total body protein, which is evidenced by a slight positive nitrogen balance when estrogens are administered. This mainly results from the growth-promoting effect of estrogen on the sexual organs, the bones, and a few other tissues of the body. The enhanced protein deposition caused by testosterone is much more general and much more powerful than that caused by estrogens.Estrogens Increase Body Metabolism and Fat Deposition
Estrogens increase the whole-body metabolic rate slightly, but only about one third as much as the increase caused by the male sex hormone testosterone. They also cause deposition of increased quantities of fat in the subcutaneous tissues. As a result, the percentage of body fat in the female body is considerably greater than that in the male body, which contains more protein. In addition to deposition of fat in the breasts and subcutaneous tissues, estrogens cause the deposition of fat in the buttocks and thighs, which is characteristic of the feminine figure.Estrogens Have Little Effect on Hair Distribution
Estrogens do not greatly affect hair distribution. However, hair does develop in the pubic region and in the axillae after puberty. Androgens formed in increased quantities by the female adrenal glands after puberty are mainly responsible for this.Effect of Estrogens on the Skin
Estrogens cause the skin to develop a texture that is soft and usually smooth, but even so, the skin of a woman is thicker than that of a child or a castrated female. Also, estrogens cause the skin to become more vascular; this is often associated with increased warmth of the skin and also promotes greater bleeding of cut surfaces than is observed in men.Effect of Estrogens on Electrolyte Balance
The chemical similarity of estrogenic hormones to adrenocortical hormones has been pointed out. Estrogens, like aldosterone and some other adrenocortical hormones, cause sodium and water retention by the kidney tubules. This effect of estrogens is normally slight and rarely of significance, but during pregnancy, the tremendous formation of estrogens by the placenta may contribute to body fluid retention, as discussed in Chapter 82.FUNCTIONS OF PROGESTERONE
Progesterone Promotes Secretory Changes in the Uterus
By far the most important function of progesterone is to promote secretory changes in the uterine endometrium during the latter half of the monthly female sexual cycle, thus preparing the uterus for implantation of the fertilized ovum. This function is discussed later in connection with the endometrial cycle of the uterus.In addition to this effect on the endometrium, progesterone decreases the frequency and intensity of uterine contractions, thereby helping to prevent expulsion of the implanted ovum.
Effect of Progesterone on the Fallopian Tubes
Progesterone also promotes increased secretion by the mucosal lining of the fallopian tubes. These secretions are necessary for nutrition of the fertilized, dividing ovum as it traverses the fallopian tube before implantation.Progesterone Promotes Development of the Breasts
Progesterone promotes development of the lobules and alveoli of the breasts, causing the alveolar cells to proliferate, enlarge, and become secretory in nature. However, progesterone does not cause the alveoli to secrete milk; as discussed in Chapter 82, milk is secreted only after the prepared breast is further stimulated by prolactin from the anterior pituitary gland.Progesterone also causes the breasts to swell. Part of this swelling is due to the secretory development in the lobules and alveoli, but part also results from increased fluid in the tissue.
MONTHLY ENDOMETRIAL CYCLE AND MENSTRUATION
Associated with the monthly cyclical production of estrogens and progesterone by the ovaries is an endometrial cycle in the lining of the uterus that operates through the following stages: (1) proliferation of the uterine endometrium; (2) development of secretory changes in the endometrium; and (3) desquamation of the endometrium, which is known as menstruation. The various phases of this endometrial cycle are shown in Figure 81-8.Proliferative Phase (Estrogen Phase) of the Endometrial Cycle, Occurring Before Ovulation
At the beginning of each monthly cycle, most of the endometrium has been desquamated by menstruation. After menstruation, only a thin layer of endometrial stroma remains and the only epithelial cells that are left are those located in the remaining deeper portions of the glands and crypts of the endometrium. Under the influence of estrogens, secreted in increasing quantities by the ovary during the first part of the monthly ovarian cycle, the stromal cells and the epithelial cells proliferate rapidly. The endometrial surface is re-epithelialized within 4 to 7 days after the beginning of menstruation.
Figure 81-8 Phases of endometrial growth and menstruation during each monthly female sexual cycle.
Then, during the next week and a half, before ovulation occurs, the endometrium increases greatly in thickness, owing to increasing numbers of stromal cells and to progressive growth of the endometrial glands and new blood vessels into the endometrium. At the time of ovulation, the endometrium is 3 to 5 millimeters thick.
The endometrial glands, especially those of the cervical region, secrete a thin, stringy mucus. The mucus strings actually align themselves along the length of the cervical canal, forming channels that help guide sperm in the proper direction from the vagina into the uterus.
Secretory Phase (Progestational Phase) of the Endometrial Cycle, Occurring After Ovulation
During most of the latter half of the monthly cycle, after ovulation has occurred, progesterone and estrogen together are secreted in large quantities by the corpus luteum. The estrogens cause slight additional cellular proliferation in the endometrium during this phase of the cycle, whereas progesterone causes marked swelling and secretory development of the endometrium. The glands increase in tortuosity; an excess of secretory substances accumulates in the glandular epithelial cells. Also, the cytoplasm of the stromal cells increases; lipid and glycogen deposits increase greatly in the stromal cells; and the blood supply to the endometrium further increases in proportion to the developing secretory activity, with the blood vessels becoming highly tortuous. At the peak of the secretory phase, about 1 week after ovulation, the endometrium has a thickness of 5 to 6 millimeters.The whole purpose of all these endometrial changes is to produce a highly secretory endometrium that contains large amounts of stored nutrients to provide appropriate conditions for implantation of a fertilized ovum during the latter half of the monthly cycle. From the time a fertilized ovum enters the uterine cavity from the fallopian tube (which occurs 3 to 4 days after ovulation) until the time the ovum implants (7 to 9 days after ovulation), the uterine secretions, called "uterine milk," provide nutrition for the early dividing ovum. Then, once the ovum implants in the endometrium, the trophoblastic cells on the surface of the implanting ovum (in the blastocyst stage) begin to digest the endometrium and absorb the endometrial stored substances, thus making great quantities of nutrients available to the early implanting embryo.
Menstruation
If the ovum is not fertilized, about 2 days before the end of the monthly cycle, the corpus luteum in the ovary suddenly involutes and the ovarian hormones (estrogens and progesterone) decrease to low levels of secretion, as shown in Figure 81-3. Menstruation follows.
Menstruation is caused by the reduction of estrogens and progesterone, especially progesterone, at the end of the monthly ovarian cycle. The first effect is decreased stimulation of the endometrial cells by these two hormones, followed rapidly by involution of the endometrium itself to about 65 percent of its previous thickness. Then, during the 24 hours preceding the onset of menstruation, the tortuous blood vessels leading to the mucosal layers of the endometrium become vasospastic, presumably because of some effect of involution, such as release of a vasoconstrictor material-possibly one of the vasoconstrictor types of prostaglandins that are present in abundance at this time.
The vasospasm, the decrease in nutrients to the endometrium, and the loss of hormonal stimulation initiate necrosis in the endometrium, especially of the blood vessels. As a result, blood at first seeps into the vascular layer of the endometrium and the hemorrhagic areas grow rapidly over a period of 24 to 36 hours. Gradually, the necrotic outer layers of the endometrium separate from the uterus at the sites of the hemorrhages until, about 48 hours after the onset of menstruation, all the superficial layers of the endometrium have desquamated. The mass of desquamated tissue and blood in the uterine cavity, plus contractile effects of prostaglandins or other substances in the decaying desquamate, all acting together, initiate uterine contractions that expel the uterine contents.
During normal menstruation, approximately 40 milliliters of blood and an additional 35 milliliters of serous fluid are lost. The menstrual fluid is normally nonclotting because a fibrinolysin is released along with the necrotic endometrial material. If excessive bleeding occurs from the uterine surface, the quantity of fibrinolysin may not be sufficient to prevent clotting. The presence of clots during menstruation is often clinical evidence of uterine pathology.
Within 4 to 7 days after menstruation starts, the loss of blood ceases because, by this time, the endometrium has become re-epithelialized.
Leukorrhea During Menstruation
During menstruation, tremendous numbers of leukocytes are released along with the necrotic material and blood. It is probable that some substance liberated by the endometrial necrosis causes this outflow of leukocytes. As a result of these leukocytes and possibly other factors, the uterus is highly resistant to infection during menstruation, even though the endometrial surfaces are denuded. This is of extreme protective value.Regulation of the Female Monthly Rhythm-Interplay Between the Ovarian and Hypothalamic-Pituitary Hormones
Now that we have presented the major cyclical changes that occur during the monthly female sexual cycle, we can attempt to explain the basic rhythmical mechanism that causes the cyclical variations.
The Hypothalamus Secretes GnRH, Which Causes the Anterior Pituitary Gland to Secrete LH and FSH
As pointed out in Chapter 74, secretion of most of the anterior pituitary hormones is controlled by "releasing hormones" formed in the hypothalamus and then transported to the anterior pituitary gland by way of the hypothalamic-hypophysial portal system. In the case of the gonadotropins, one releasing hormone, GnRH, is important. This hormone has been purified and has been found to be a decapeptide with the following formula:

Hypothalamic Centers for Release of GnRH
The neuronal activity that causes pulsatile release of GnRH occurs primarily in the mediobasal hypothalamus, especially in the arcuate nuclei of this area. Therefore, it is believed that these arcuate nuclei control most female sexual activity, although neurons located in the preoptic area of the anterior hypothalamus also secrete GnRH in moderate amounts. Multiple neuronal centers in the higher brain's "limbic" system (the system for psychic control) transmit signals into the arcuate nuclei to modify both the intensity of GnRH release and the frequency of the pulses, thus providing a partial explanation of why psychic factors often modify female sexual function.Negative Feedback Effects of Estrogen and Progesterone to Decrease LH and FSH Secretion
Estrogen in small amounts has a strong effect to inhibit the production of both LH and FSH. Also, when progesterone is available, the inhibitory effect of estrogen is multiplied, even though progesterone by itself has little effect (Figure 81-10).These feedback effects seem to operate mainly on the anterior pituitary gland directly, but they also operate to a lesser extent on the hypothalamus to decrease secretion of GnRH, especially by altering the frequency of the GnRH pulses.
Inhibin from the Corpus Luteum Inhibits FSH and LH Secretion
In addition to the feedback effects of estrogen and progesterone, other hormones seem to be involved, especially inhibin, which is secreted along with the steroid sex hormones by the granulosa cells of the ovarian corpus luteum in the same way that Sertoli cells secrete inhibin in the male testes (see Figure 81-10). This hormone has the same effect in the female as in the male-inhibiting the secretion of FSH and, to a lesser extent, LH by the anterior pituitary gland. Therefore, it is believed that inhibin might be especially important in causing the decrease in secretion of FSH and LH at the end of the monthly female sexual cycle.Positive Feedback Effect of Estrogen Before Ovulation-The Preovulatory LH Surge
For reasons not completely understood, the anterior pituitary gland secretes greatly increased amounts of LH for 1 to 2 days beginning 24 to 48 hours before ovulation. This effect is demonstrated in Figure 81-3. The figure shows a much smaller preovulatory surge of FSH as well.Experiments have shown that infusion of estrogen into a female above a critical rate for 2 to 3 days during the latter part of the first half of the ovarian cycle will cause rapidly accelerating growth of the ovarian follicles, as well as rapidly accelerating secretion of ovarian estrogens. During this period, secretions of both FSH and LH by the anterior pituitary gland are at first slightly suppressed. Then secretion of LH increases abruptly sixfold to eightfold, and secretion of FSH increases about twofold. The greatly increased secretion of LH causes ovulation to occur.
The cause of this abrupt surge in LH secretion is not known. However, several possible explanations are as follows: (1) It has been suggested that estrogen at this point in the cycle has a peculiar positive feedback effect of stimulating pituitary secretion of LH and, to a lesser extent, FSH (see Figure 81-10); this is in sharp contrast to its normal negative feedback effect that occurs during the remainder of the female monthly cycle. (2) The granulosa cells of the follicles begin to secrete small but increasing quantities of progesterone a day or so before the preovulatory LH surge, and it has been suggested that this might be the factor that stimulates the excess LH secretion.
Without this normal preovulatory surge of LH, ovulation will not occur.
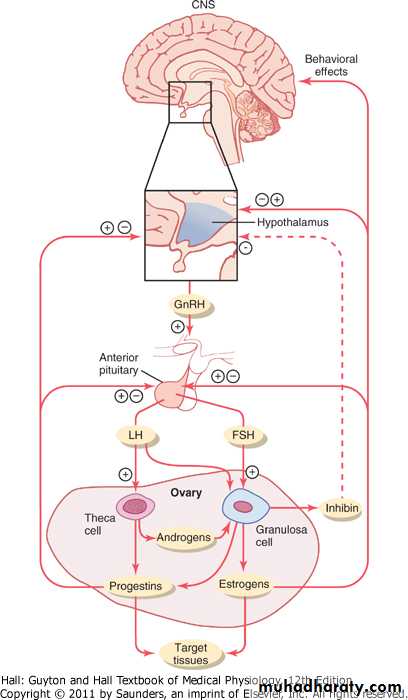
Figure 81-10 Feedback regulation of the hypothalamic-pituitary-ovarian axis in females. Stimulatory effects are shown by + and negative feedback inhibitory effects are shown by -. Estrogens and progestins exert both negative and positive feedback effects on the anterior pituitary and hypothalamus depending on the stage of the ovarian cycle. Inhibin has a negative feedback effect on the anterior pituitary. FSH, follicle-stimulating hormone; GnRH, gonadotropin-releasing hormone; LH, luteinizing hormone.