
BY
Dr. Dhafer Abdullah Farhan
Ph. D. Cancer research
Today We are here
List of abbreviations

Introduction
Adaptive immune responses have been subdivided into:Humoral (antibody- mediated) immune responses.
Cellular (cell-mediated) immune responses.
* Host defenses against extracellular infections are mediated by:
AntibodyComplement
Macrophages
* Intercellular infections are mediates by CMI
* CMI are responsible for:
- Resistance to intracellular pathogens- Resistance to fungal and protozoal infections
- Resistance to tumors
Cell Mediated Immunity
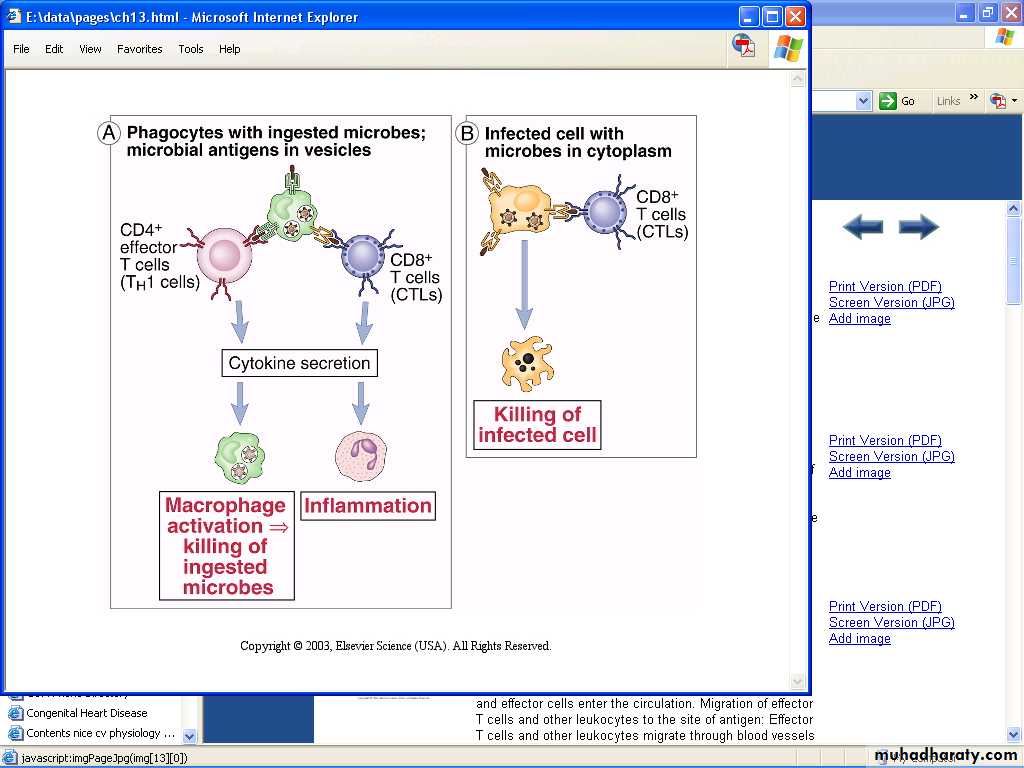
Many microbes have developed mechanisms that enable them to survive and even replicate within phagocytes, so the innate immunity is unable to eradicate infections by such microbes.In CMI against phagocytosed microbes, the specificity of the response is due to T cells – but the actual effector function is mediated by the phagocytes.
Cell-mediated immunity
CMI is an immune response that does not involve antibodies but rather involves :the activation of macrophages and NK-cells.
The production of antigen-specific cytotoxic T-lymphocytes.
The release of various cytokines in response to an antigen.
It occurs inside infected cells and is mediated by T lymphocytes.
The pathogen's antigens are expressed on the cell surface or on an antigen-presenting cell. Helper T cells release cytokines that help activated T cells bind to the infected cells’ MHC-antigen complex and differentiate the T cell into a cytotoxic T cell. The infected cell then undergoes lysis.Cellular immunity protects the body by:
• Activating antigen-specific cytotoxic T-lymphocytes (CTLs) that are able to destroy body cells displaying epitopes of foreign antigen on their surface, such as virus-infected cells, cells with intracellular bacteria, and cancer cells displaying tumor antigens;• Activating macrophages and NK cells, enabling them to destroy intracellular pathogens.
• Stimulating cells to secrete a variety of cytokines that influence the function of other cells involved in adaptive immune responses and innate immune responses.
Cell Mediated Immunity
Specialized lymphocytes, mostly T cells, respond to intracellular antigens.
After differentiating in the thymus, T cells migrate to lymphoid tissue.
T cells differentiate into effector T cells when stimulated by an Antigen.
Some effector T cells become memory cells.
Helper T Cell (CD4, TH).
TH1 Activate cells related to cell-mediated immunity.TH2 Activate B cells to produce IgM, and IgE
Cytotoxic T Cells (CD8, TC)
Destroy target cells with perforin
Delayed Hypersensitivity T Cells (TD) (CD4):
Associated with allergic reaction, transplant rejection, and tuberculin skin test.
Suppressor T cells (TS) (CD8)
Turn off immune response when Ag no longer present
Characters Of CMI
Cellular immune response is mediated by:- Subpopulation of T-lymphocytes
- Macrophages and their products
What Does the T Cell Receptor (TCR) Recognize?
Only fragments of proteins (peptides) associated with MHC molecules on surface of cells.Helper T cells (TH) recognize peptide associated with MHC class II molecules.
Cytotoxic T cells (Tc) recognize peptide associated with MHC class I molecules

Cell-mediated cytotoxicity
It is an essential defense against intracellular pathogens, including viruses, some bacteria and parasites, tumor cells and even cells of the body.In addition, the process is important in the destruction of allogeneic tissue grafts.
Several types of cells can execute this activity including
T-cytotoxic (Tc) cells, NK-cells and sometimes myeloid cells.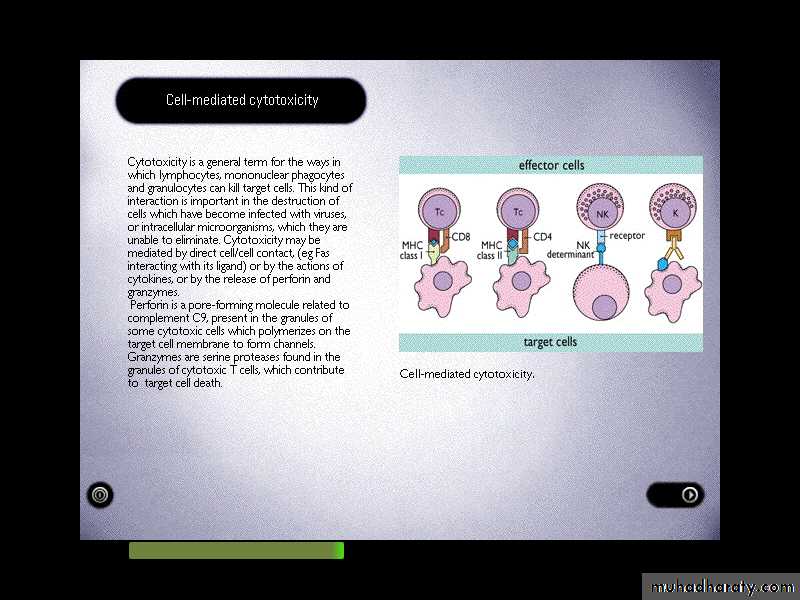
Cellular immunity protects the body through:
• T-cell mediated immunity activating antigen-specific cytotoxic T cells that are able to induce apoptosis in body cells displaying epitopes of foreign antigen on their surface, such as virus-infected cells, cells with intracellular bacteria, and cancer cells displaying tumor antigens.• Macrophage and natural killer cell action: enabling the destruction of pathogens via recognition and secretion of cytotoxic granules (for natural killer cells) and phagocytosis (for macrophages) and
• Stimulating cells to secrete a variety of cytokines that influence the function of other cells involved in adaptive immune responses and innate immune responses.
Tc-cells recognize specific Ag presented by MHC-I.
Most Tc are CD8+ve, but about 10% of MHC-restricted cytotoxic T-cells are CD4+ve & recognize Ag presented on MHC-II.Tc-cells and NK-cells act in a complementary
way to protect the body.The TCRs and CD8 molecules on the surface of naive T8-lymphocytes are designed to recognize peptide epitopes bound to MHC-I molecules on antigen-presenting cells or APCs .
The TCRs and CD8 molecules on the surface of cytotoxic T-lymphocytes (CTLs) are designed to recognize peptide epitopes bound to MHC-I molecules on infected cells and tumor cells.
NK-cells
• Recognize cells which fail to express MHCI molecules.• They can also bind to Ab already attached to Ag an a target cell, using their Fc-R (CD16), in a what is known as AB-dependent cell-mediated cytotoxicity (ADCC).
Several viruses have evolved mechanisms to avoid recognition by Tc cell.
• They reduce the expression of MHC molecules.
• Produce proteins which divert MHC molecules out of the endoplasmic reticulum.So NK-cells specifically recognize cells which have lost their MHC - I molecules.
Mechanisms of cell-mediatedcytotoxicity
Tc, NK- and K-cells use variety of different mechanisms to kill their targets, including: * direct cell-cell signaling via surface molecules. * indirect signaling via cytokines.
in addition many CD8 Tc and large granular lymphocytes (NK and K-cells) have granules which contain proteins that damage target cells if they are released directly against the target cell plasma membrane.
Tc cells signal to their targets using members of TNF-R group of molecules.
These include Fas (CD95) and the type-1 TNF-R (TNFR-1) which are widely distributed in the body.CD8+ T cell can kill the infected cells is via Fas/Fas L interactions. Activated CD8+ T cells express Fas L on the cell surface, which binds to its receptor, Fas, on the surface of the target cell.
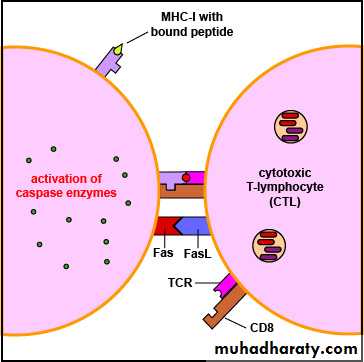
A cytotoxicity which may be signaled via:
Fas or a TNF-R on a target cell.
This binding causes the Fas molecules on the surface of the target cell to trimerise, which pulls together signalling molecules. These signalling molecules result in the activation of the caspase cascade, which also results in apoptosis of the target cell.
** Most CD8 TC and NK-cells (and macrophages) have vesicles containing TNF and lymphotoxin which can be released onto a target cells.
TNF acts in a very similar way to the Fas-ligand. It causes trimerization of the TNFR-1 so that the receptor associates with adaptor proteins which recruit caspases.
Both TNFR-1 and Fas containing intracytoplasmic domains (death-domains) which are found on a number of proteins involved in cell survival.
*** The specific granules of NK-cells and Tc contain several proteins, including perforin and granzymes (granules associated enzymes). After binding to its target, the Tc cells directs its granules towards the membrane a joining the target. Then, in a Ca-dependent phase, the granule contents are discharge into the cleft between the two cells.
Perforin is a pore-forming proteins and in the presence of Ca , the perforin bind to the target cell membrane and polymerize to form transmembrane channels. Tc-protected from auto-destruction with this protein by a proteoglycan (chondroitin-sulphate A) which may bind to and inactivate the perforin, thus Tc-survives and can continue kill further targets.
Granzymes are collection of serine estrases (enzymes) which are also released upon granule exocytosis and become active after release. Some of these enzyme trigger apoptosis and DNA-degradation.
Both perforin and granzymes act synergistically; the granzymes enter the target cells via pore created by perforin.
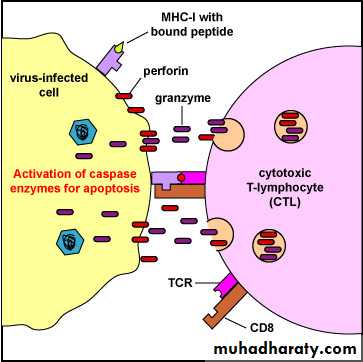
Cytotoxic T-lymphocyte (CTL)-Induced Apoptosis of a virus-Infected Cell. Binding of the CTL to the infected cell triggers the CTL to release pore-forming proteins called perforins, proteolytic enzymes called granzymes, and chemokines. Granzymes pass through the pores and activate the caspase enzymes that lead to apoptosis of the infected cell by means of destruction of its structural cytoskeleton proteins and by chromosomal degradation. As a result, the cell breaks into fragments that are subsequently removed by phagocytes.
To summarize the lymphoid cytotoxic effectors:
CD8 Tc use both Fas- L and granule release to kill their targets.CD4 Tc-cells use principally FasL, and NK-cells use primarily their granules.
TNF may contribute to the cytotoxic damage produced by any of these cells.
*Cytotoxicity may be triggered specifically to a target by ADCC or may involve a range of non-specific toxic mediators. Macrophage and neutrophil express Fc-gR1&2 which allow them to engage their targets by ADCC.`
A number of non-lymphoid cells may be cytotoxic to other cells
** Macrophages and neutrophils granules which contain neutrophil defensins, lysosomal enzymes and cytostatic proteins. If the phagocyte fails to internalize its target, then these mediators may be released into the extracellular environment and contribute to localized cell damage ((frustrated phagocytosis)), the effect of these mediators tends to produce necrosis and inflammation.
A number of non-lymphoid cells may be cytotoxic to other cells
*** Activated macrophage secrete TNF which can induce apoptosis in a similar way to lymphoid cytotoxic effectors.Eosinophils are characterized by their granules. Eosinophils are only weakly phagocytic, they ingest some bacteria following activation but are less efficient than neutrophils at intracellular killing.
Their major function appears to be the secretion of various toxic granules. They are therefore effective for the extracellular killing of microorganisms, particularly large parasites such as Schistosoma.
The components of the eosinophil granule include major basic protein (MBP), eosinophil peroxidase (EPO) and eosinophil cationic protein (ECP).
Other molecules produced by eosinophils are lysophospholipase and eosinophil-derived neurotoxin (EDN), which are also a ribonuclease but with strong neurotoxic activity. Thus, killing may involve contact-dependent degranulation or may simply require deposition of toxins within the local tissue.

THANK YOU
