Cells physiology
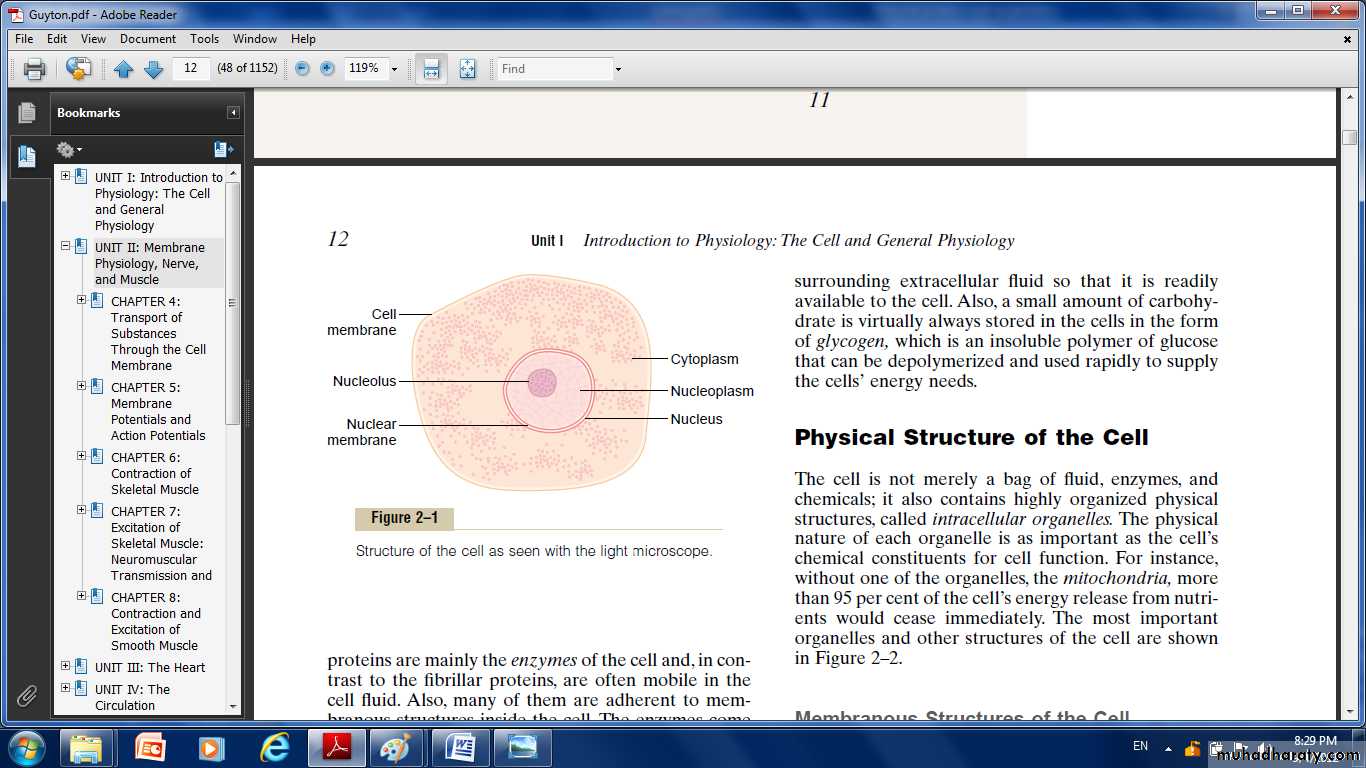
Organization of the CellA typical cell formed by two major parts the nucleus and the cytoplasm. The nucleus is separated from the cytoplasm by a nuclear membrane, and the cytoplasm is separated from the surrounding fluids by a cell membrane. The different substances that make up the cell are collectively called protoplasm. Protoplasm is composed mainly of five basic substances: water, electrolytes, proteins, lipids, and carbohydrates.
Membranous Structures of the Cell
Most organelles of the cell are covered by membranes composed primarily of lipids and proteins. The lipids of the membranes provide a barrier that impedes the movement of water and water-soluble substances from one cell compartment to another because water is not soluble in lipids. Protein molecules in the membrane often do penetrate all the way through the membrane, thus providing specialized pathways, often organized into actual pores, for passage of specific substances through the membrane, other membrane proteins are enzymes that catalyze a multitude of different chemical reactions.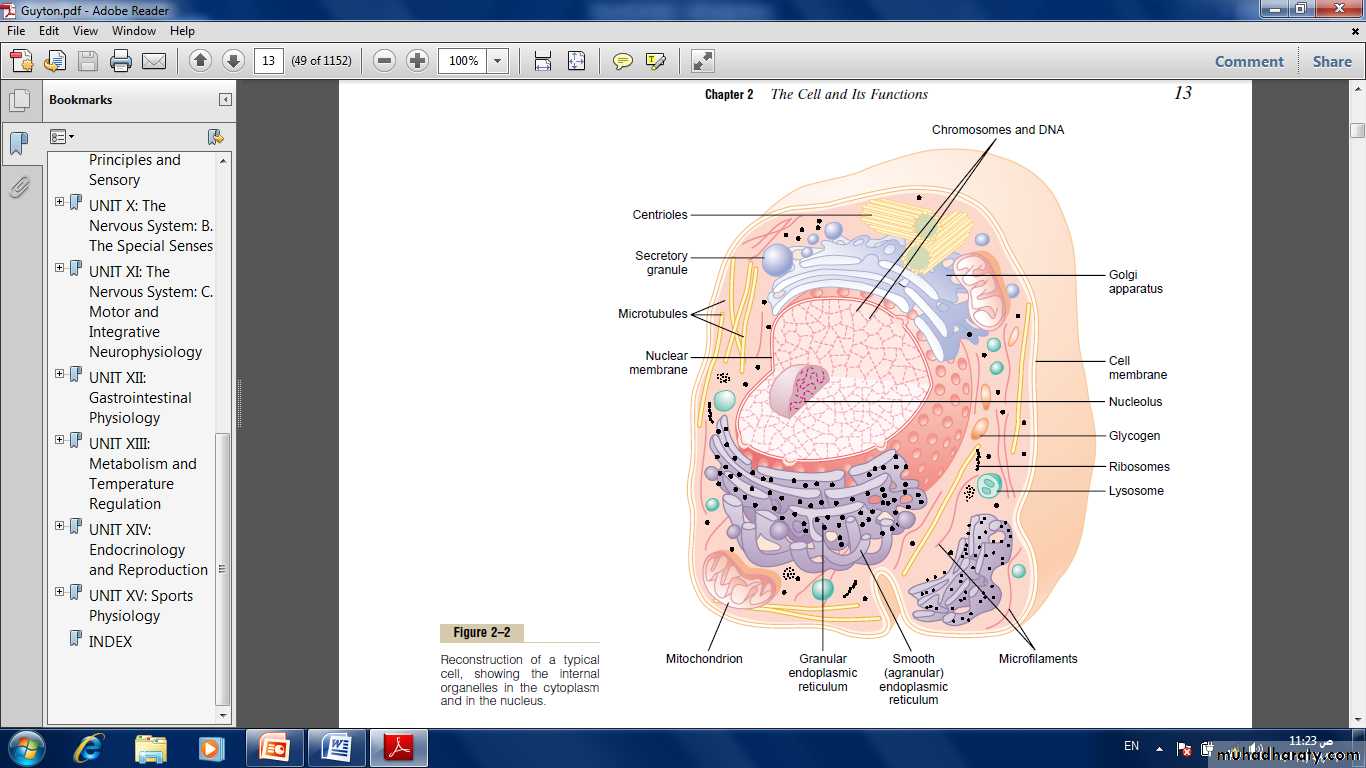
Fig: Reconstruction of a typical cell, showing the internal organelles in the cytoplasm and in the nucleus.
The cell membrane (also called the plasma membrane),which envelops the cell, is a thin, pliable, elastic structure only 7.5 to 10 nanometers thick. It is composed almost entirely of proteins and lipids. The approximate composition is proteins 55%; phospholipids 25%; cholesterol 13%; other lipids 4%; and carbohydrates 3%.
Lipid Barrier of the Cell Membrane Impedes Water Penetration:
The basic structure of cell membrane is a lipid bilayer, which is a thin, double-layered film of lipids—each layer only one molecule thick—that is continuous over the entire cell surface. Interspersed in this lipid film are large globular protein molecules. The basic lipid bilayer is composed of phospholipid molecules. One end of each phospholipid molecule is soluble in water; that is, it is hydrophilic. The other end is soluble only in fats; that is, it is hydrophobic. The lipid layer in the
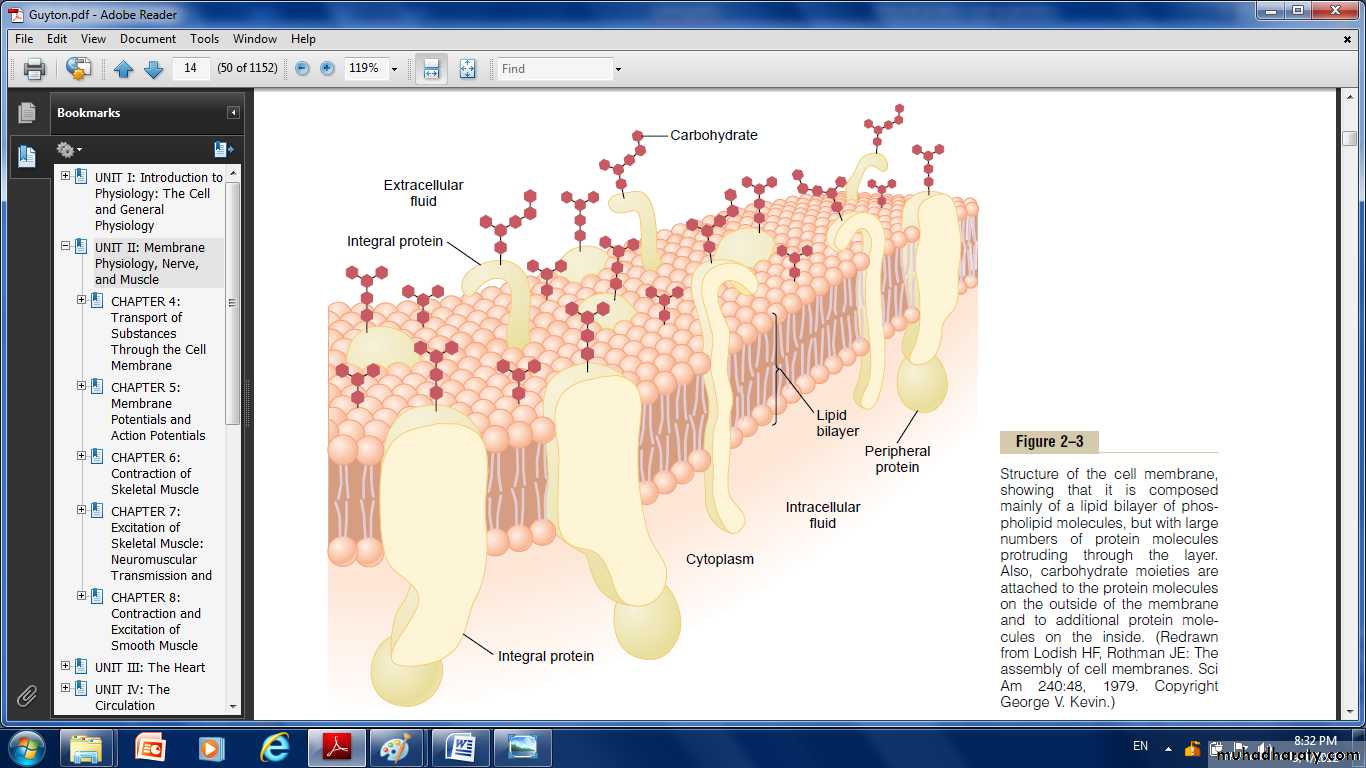
Fig: structure of the cell membrane, showing that it is composed mainly of a lipid bilayer of phospholipid molecules.but with large number of protein molecules protruding through the layer. Also carbohydrate molecules are attached to the protein molecules on the outside of the membrane and to additional protein molecule to the inside.
middle of the membrane is impermeable to the usual water-soluble substances, such as ions, glucose, and urea.
Conversely, fat-soluble substances, such as oxygen, carbon dioxide, and alcohol, can penetrate this portion of the membrane with ease.
Cell Membrane Proteins.
There are globular masses floating in the lipid bilayer. These are membrane proteins, most of which are glycoproteins. Two types of proteins are present:
Integral proteins:
They protrude all the way through the membrane, Many of the integral proteins provide structural channels (or pores) through which water molecules and water-soluble substances, especially ions, can diffuse between the extracellular and intracellular fluids. These protein channels also have selective properties that allow preferential diffusion of some substances over others. Integral membrane proteins can also serve as a receptors for water-soluble chemicals, such as peptide hormones, that do not easily penetrate the cell membrane. Still others act as enzymes.
Carrier proteins:
They transport substances that otherwise could not penetrate the lipid bilayer . Sometimes these even transport substances in the direction opposite to their natural direction of diffusion, which is called “active transport.”
Membrane Carbohydrates—The Cell “Glycocalyx.”
Membrane carbohydrates occur almost invariably in combination with proteins or lipids in the form of glycoprotein or glycolipids, the entire outside surface of the cell often has a loose carbohydrate coat called the glycocalyx. The carbohydrate moieties attached to the outer surface of the cell have several important functions:
(1) Many of them have a negative electrical charge, which gives most cells an overall negative surface charge that repels other negative objects. (2) The glycocalyx of some cells attaches to the glycocalyx of other cells, thus attaching cells to one another.
(3) Many of the carbohydrates act as receptor substances for binding hormones, such as insulin.
(4) Some carbohydrate moieties enter into immune reactions.
Cytoplasm and Its Organelles
The cytoplasm is filled with both minute and large dispersed particles and organelles. There are five important organelles: the endoplasmic reticulum, Golgi apparatus, mitochondria, lysosomes and peroxisomes.
Endoplasmic Reticulum
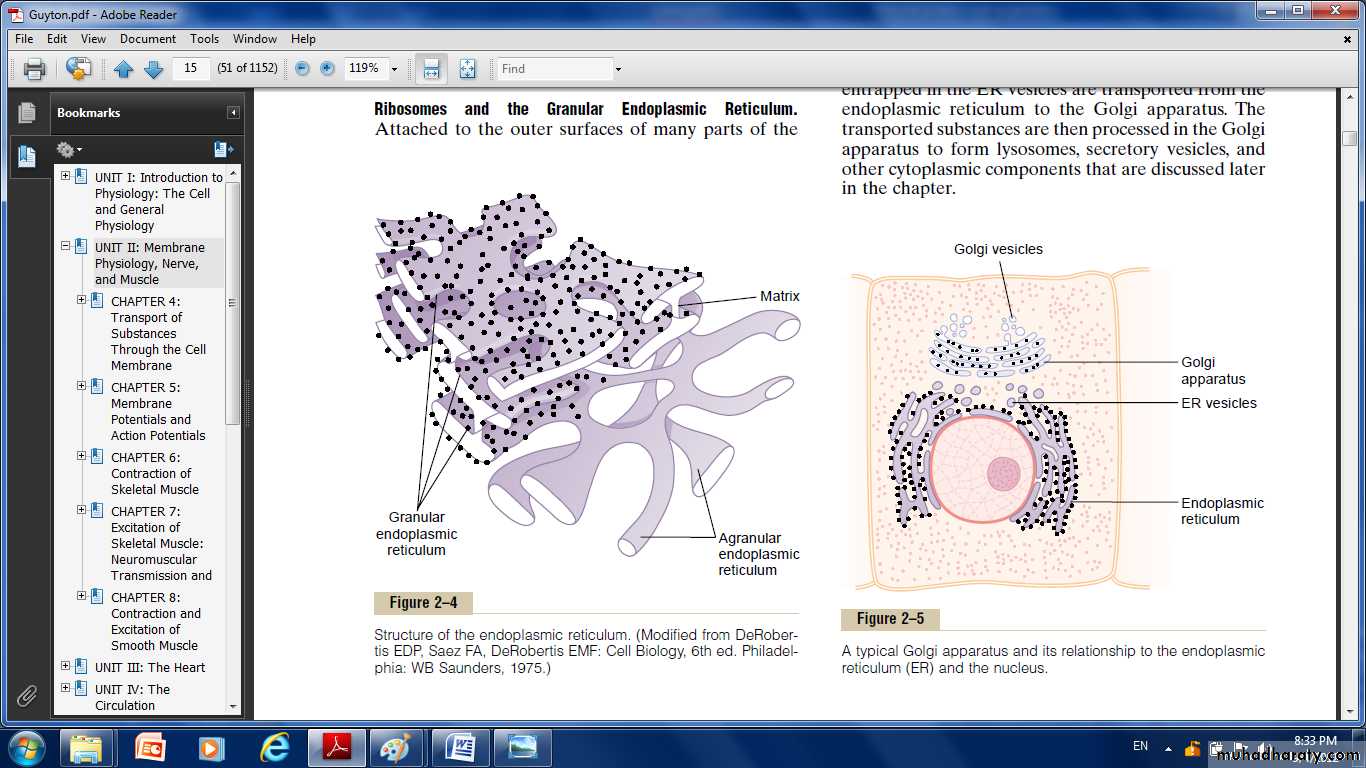
Fig: Structure of endoplasmic reticulum
It's a network of tubular and flat vesicular structures in the cytoplasm, their walls are constructed of lipid bilayer membranes that contain large amounts of proteins.
The vast surface area of this reticulum and the multiple enzyme systems attached to its membranes provide machinery for a major share of the metabolic functions of the cell.
Ribosomes and the Granular Endoplasmic Reticulum. Attached to the outer surfaces of many parts of the endoplasmic reticulum are large numbers of minute granular particles called ribosomes. Where these are present, the reticulum is called the granular endoplasmic
Reticulum, they function to synthesize new protein molecules in the cell.
A granular Endoplasmic Reticulum.
Part of the endoplasmic reticulum has no attached ribosomes. This part is called the a granular, or smooth, endoplasmic reticulum. The a granular reticulum functions is synthesis of lipid substances.
Golgi Apparatus:
Is closely related to the endoplasmic reticulum. It has membranes similar to those of the a granular endoplasmic reticulum. It usually is composed
of four or more stacked layers of thin, flat, enclosed vesicles lying near one side of the nucleus. This apparatus is prominent in secretory cells, where it is located on the side of the cell from which the secretory substances are extruded. The Golgi apparatus functions in association with the endoplasmic reticulum, small “transport vesicles” continually pinch off from the endoplasmic reticulum and shortly thereafter fuse with the Golgi apparatus. In this way, substances entrapped in the ER vesicles are transported from the endoplasmic reticulum to the Golgi apparatus. The transported substances are then processed in the Golgi apparatus to form lysosomes, secretory vesicles and other cytoplasmic components.
Lysosomes: Lysosomes are vesicular organelles that form by breaking off from the Golgi apparatus and then dispersing throughout the cytoplasm. The lysosomes provide an intracellular digestive system that allows the cell to digest:
(1) Damaged cellular structures.
(2) Food particles that have been ingested by the cell.
(3) Unwanted matter such as bacteria.
It is surrounded by a typical lipid bilayer membrane and is filled with large numbers of small granules, which are protein aggregates of as many as 40 different hydrolase (digestive) enzymes. A hydrolytic enzyme is capable of splitting an organic compound into two or more parts.
Peroxisomes: Peroxisomes are similar physically to lysosomes, but they are different in two important ways:
First, they are believed to be formed by self-replication (or perhaps by budding off from the smooth endoplasmic reticulum) rather than from the Golgi apparatus.
Second, they contain oxidases rather than hydrolases. Several of the oxidases are capable of combining oxygen with hydrogen ions derived from different intracellular chemicals to form hydrogen peroxide (H2O2). Hydrogen peroxide is a highly oxidizing substance and is used in association with catalase, another oxidase enzyme present in large quantities in peroxisomes, to oxidize many substances that might otherwise be poisonous to the cell.
Secretory Vesicles:
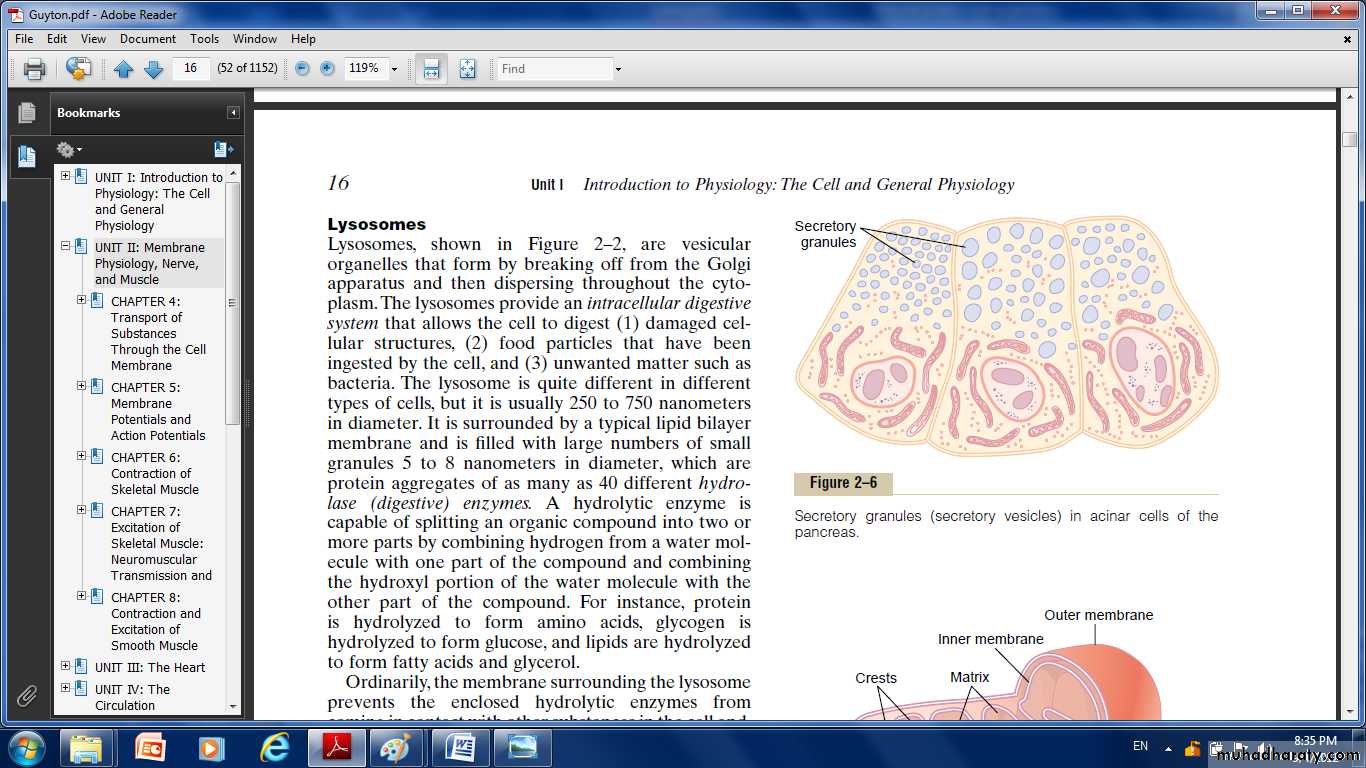
Fig: Secretory granules (secretory vesicles) in acinar cells of the pancrease.
One of the important functions of many cells is secretion of special chemical substances. Almost all such secretory substances are formed by the endoplasmic reticulum–Golgi apparatus system and are then released from the Golgi apparatus into the cytoplasm in the form of storage vesicles called secretory vesicles or secretory granules.Mitochondria:
The mitochondria also called the “power houses” of the cell. Without them, cells would be unable to extract enough energy from the nutrients, and essentially all cellular functions would cease. Mitochondria are present in all areas of each cell’s cytoplasm, but the total number per cell
varies from less than a hundred up to several thousand, depending on the amount of energy required by the cell. Further, the mitochondria are concentrated in those portions of the cell that are responsible for the major share of its energy metabolism. They are also variable in size and
shape. it composed mainly of two lipid bilayer –protein membranes:
an outer membrane and an inner membrane. Many infoldings of the inner membrane form shelves onto which oxidative enzymes are attached; the inner cavity of the mitochondrion is filled with a matrix that contains large quantities of dissolved enzymes that are necessary for extracting energy from nutrients.
These enzymes operate in association with the oxidative enzymes on the shelves to cause oxidation of the nutrients, thereby forming carbon dioxide and water and at the same time releasing energy. The liberated energy is used to synthesize a “high-energy” substance called adenosine triphosphate (ATP).ATP is then transported out of the mitochondrion, and it diffuses throughout the cell to release its own energy wherever it is
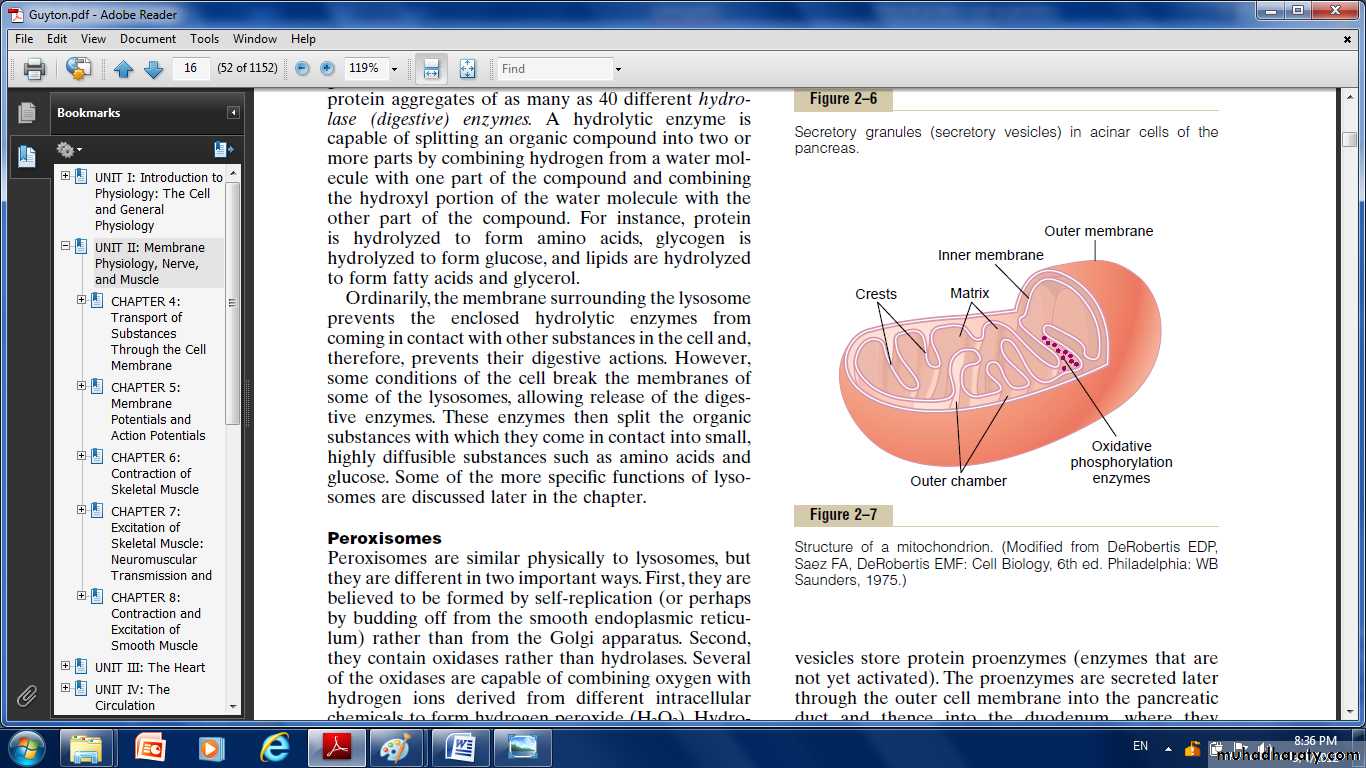
Fig. Structure of the mitochondrid.
needed for performing cellular function. Mitochondria are self replicative, which means that one mitochondrion can form a second one, a third one, and so on, whenever there is a need in the cell for increased amounts of ATP. Indeed, the mitochondria contain DNA similar to that found in the cell nucleus. The DNA of the mitochondrion plays a role in controlling replication of the mitochondrion itself.Filament and Tubular Structures of the Cell
The fibrillar proteins of the cell are usually organized into filaments or tubules. These originate as precursor protein molecules synthesized by ribosomes in the cytoplasm. The precursor molecules then polymerize to form filaments.
Nucleus: The nucleus is the control center of the cell. Briefly, the nucleus contains large quantities of DNA, which are the genes. The genes determine the characteristics of the cell’s proteins, including the structural
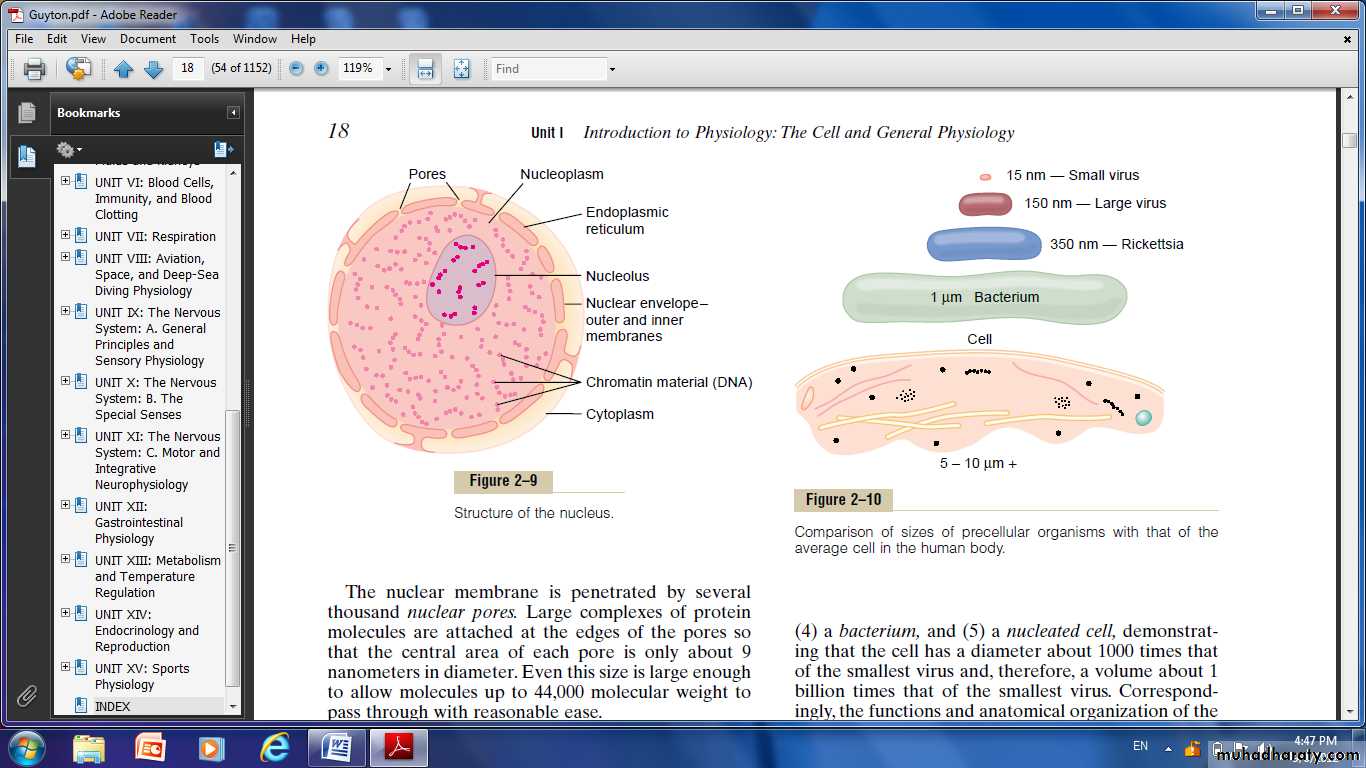
Fig. Structure of the nucleus
Proteins and the intracellular enzymes that control cytoplasmic and nuclear activities. The genes also control and promote reproduction of the cell itself. The genes first reproduce to give two identical sets of genes; then the cell splits by a special process called mitosis to form two daughter cells, each of which receives one of the two sets of DNA genes.
Nucleoli and Formation of Ribosomes
The nuclei of most cells contain one or more highly staining structures called nucleoli. The nucleolus does not have a limiting membrane. Instead, it is simply an accumulation of large amounts of RNA and proteins of the types found in ribosomes. The nucleolus becomes considerably enlarged when the cell is actively synthesizing proteins. Formation of the nucleoli (and of the ribosomes in the cytoplasm outside the nucleus) begins in the nucleus.
Transport of Substances Through the Cell Membrane “Diffusion” Versus “Active Transport.”
Transport through the cell membrane, either directly through the lipid bilayer or through the proteins, occurs by one of two basic processes:
Diffusion.
Active transport.
Diffusion: means random molecular movement of substances molecule by molecule, either through intermolecular spaces in the membrane or in combination with a carrier protein.
Active transport: means movement of ions or other substances across the membrane in combination with a carrier protein in such a
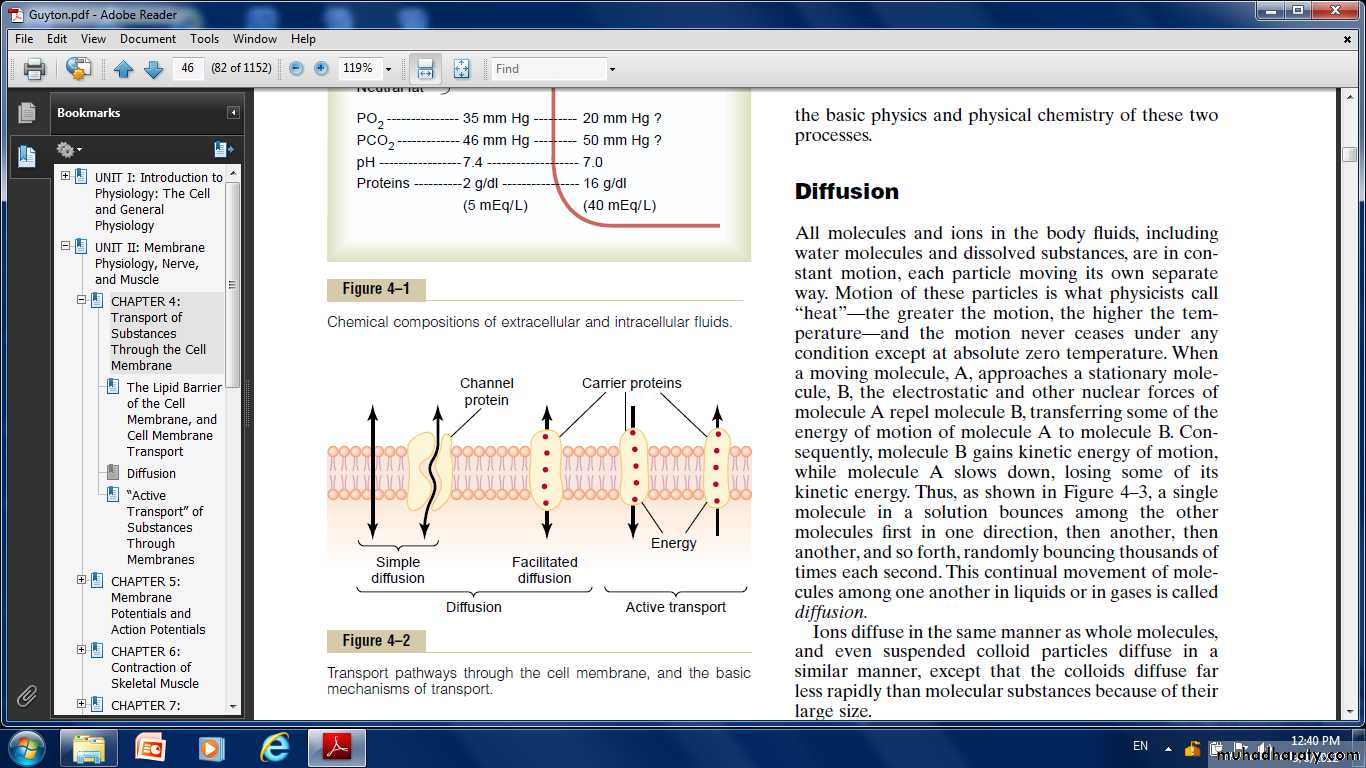
way that the carrier protein causes the substance to move against an energy gradient, such as from a low-concentration state to a high-concentration state. This movement requires an additional source of energy besides kinetic energy.
Diffusion Through the Cell Membrane
Simple diffusion and facilitated diffusion.
Simple diffusion: Means that kinetic movement of molecules or ions occurs through a membrane opening or through intermolecular spaces without any interaction with carrier proteins in the membrane. The rate of diffusion is determined by the amount of substance available, the velocity of kinetic motion, and the number and sizes of openings in the membrane through which the molecules or ions can move. Simple diffusion can occur through the cell membrane by two pathways:
(1) Through the interstices of the lipid bilayer if the diffusing substance is lipid soluble.
(2) Through watery channels that penetrate all the way through some of the large transport proteins.
Facilitated diffusion:
Requires interaction of a carrier protein. The carrier protein aids passage of the molecules or ions through the membrane by binding chemically with them and shuttling them through the membrane in this form.
Diffusion of Lipid-Soluble Substances Through the Lipid Bilayer.
One of the most important factors that determines how rapidly a substance diffuses through the lipid bilayer is the lipid solubility of the substance. For instance, the lipid solubilities of oxygen, nitrogen, carbon dioxide, and alcohols are high, so that all these can dissolve directly in the lipid bilayer and diffuse through the cell membrane in the same manner that diffusion of water solutes occurs in a watery solution.
Diffusion of Water and Other Lipid-Insoluble Molecules Through Protein Channels.
Even though water is highly insoluble in the membrane lipids, it readily passes through channels in protein molecules that penetrate all the way through the membrane. Other lipid-insoluble molecules can pass through the protein pore channels in the same way as water molecules if they are water soluble and small enough.
Diffusion Through Protein Channels, and “Gating” of These Channels:
Substances can move by simple diffusion directly along these channels from one side of the membrane to the other. The protein channels are distinguished by two important characteristics:
They are often selectively permeable to certain substances.
(2) Many of the channels can be opened or closed by gates.
Selective Permeability of Protein Channels.
Many of the protein channels are highly selective for transport of one or more specific ions or molecules. This results from the characteristics of the channel itself, such as its diameter, its shape, and the nature of the electrical charges and chemical bonds along its inside surfaces. To give an example, one of the most important of the protein channels:
Sodium channels: They are only 0.3 by 0.5 nanometer in diameter, the inner surfaces of this channel are strongly negatively charged. These strong negative charges can pull small dehydrated sodium ions into these channels, actually pulling the sodium ions away from their hydrating water molecules. Thus, the sodium channel is specifically selective for passage of sodium ions.
Potassium channels: These channels are slightly smaller than the sodium channels, only 0.3 by 0.3 nanometer, but they are not negatively charged, and their chemical bonds are different. Therefore, no strong attractive force is pulling ions into the channels, and the potassium ions are not pulled away from the water molecules that hydrate them.
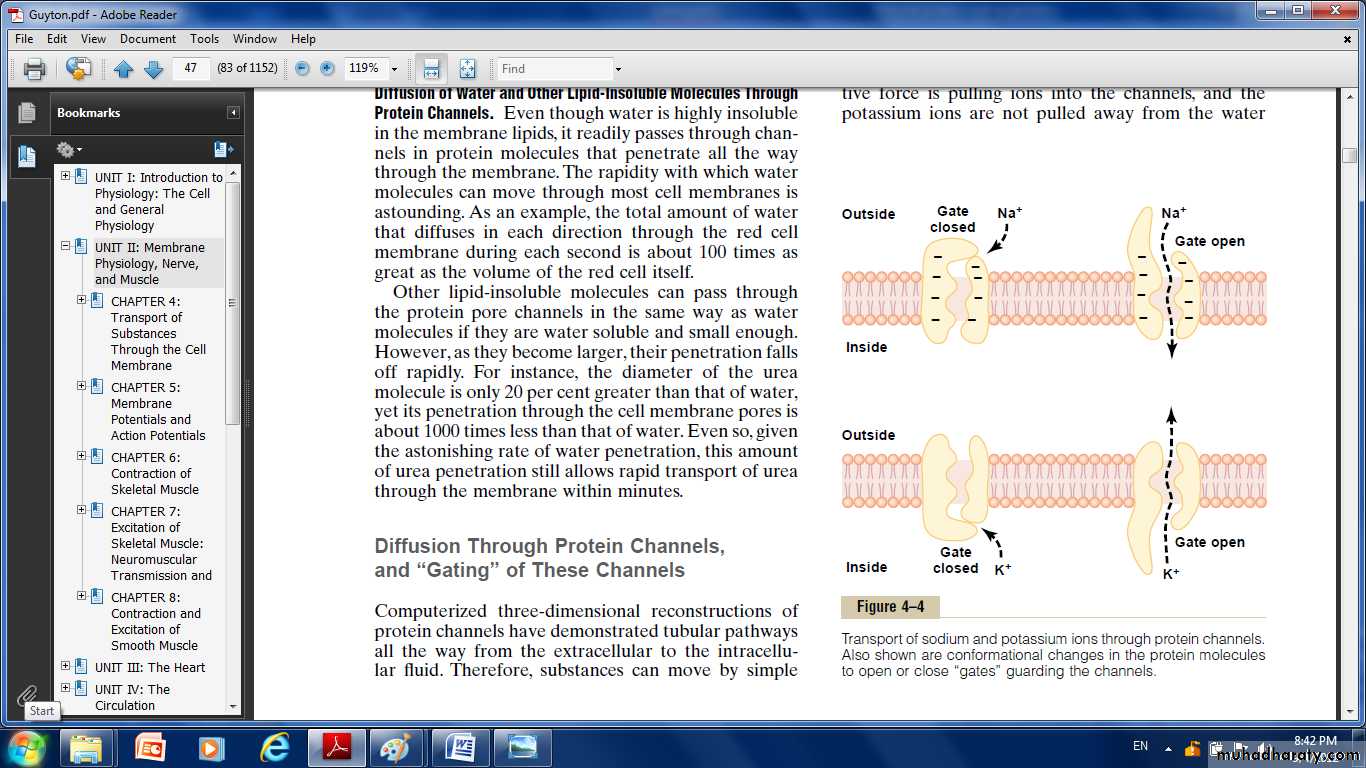
The hydrated form of the potassium ion is considerably smaller than the hydrated form of sodium because the sodium ion attracts far more water molecules than does potassium. Therefore, the smaller hydrated potassium ions can pass easily through this small channel, whereas the larger hydrated sodium ions are rejected, thus providing selective permeability for a specific ion.
Gating of Protein Channels.
Gating of protein channels provides a means of controlling ion permeability of the channels. It is believed that some of the gates are actual gate like extensions of the transport protein molecule, which can close the opening of the channel or can be lifted away from the opening by a conformational change in the shape of the protein molecule itself. The opening and closing of gates are controlled in two principal ways:
Voltage gating.
when there is a strong negative charge on the inside of the cell membrane, this presumably could cause the outside sodium gates to remain tightly closed; conversely, when the inside of the membrane loses its negative charge, these gates would open suddenly and allow tremendous quantities of sodium to pass inward through the sodium pores. This is the basic mechanism for eliciting action potentials in nerves that are responsible for nerve signals. While the potassium gates are on the intracellular ends of the potassium channels, and they open when the inside of the cell membrane becomes positively charged. The opening of these gates is partly responsible for terminating the action potential.
2. Chemical (ligand) gating.
Some protein channel gates are opened by the binding of a chemical substance (a ligand) with the protein; this causes a conformational or chemical bonding change in the protein molecule that opens or closes the gate. This is called chemical gating or ligand gating. One of the most important instances of chemical gating is the effect of acetylcholine on the so-called acetylcholine channel. Among the most important substances that cross cell membranes by facilitated diffusion are glucose and most of the amino acids. In the case of glucose, the carrier molecule has been discovered, and it has a molecular weight of about 45,000; it can also transport several other monosaccharides that have structures similar to that of glucose, including galactose. Also, insulin can increase the rate of facilitated diffusion of glucose as much as 10-fold to 20-fold. This is the principal mechanism by which insulin controls glucose use in the body.
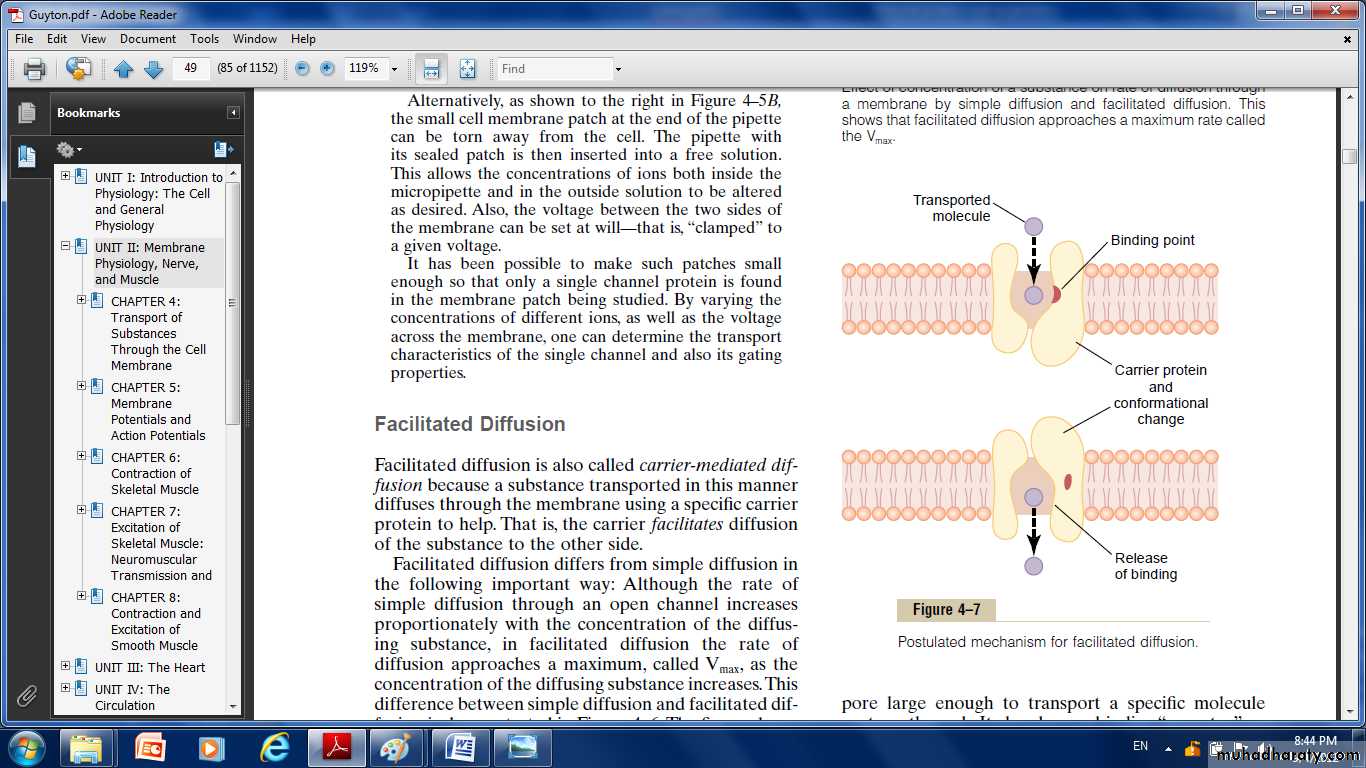
Factors That Affect Net Rate of Diffusion:
Concentration Difference : The rate at which the substance diffuses inward is proportional to the concentration of molecules on theoutside, because this concentration determines how many molecules strike the outside of the membrane each second.Conversely, the rate at which molecules diffuse outward is proportional to their concentration inside the membrane. Therefore, the rate of net diffusion into the cell is proportional to the concentration on the outside minus the concentration on the inside, or: Net diffusion μ (Co - Ci) in which Co is concentration outside and Ci is concentration inside.
Membrane Electrical Potential. If an electrical potential is applied across the membrane, the electrical charges of the ions cause them to move through the membrane even though no concentration difference exists to cause movement.
Effect of a Pressure Difference Across the Membrane. Considerable pressure difference develops between the two sides of a diffusible membrane. This occurs, for instance, at the blood capillary membrane in all tissues of the body. The pressure is about 20 mm Hg greater inside the capillary than outside.
“Active Transport” of Substances Through Membranes: At times, a large concentration of a substance is required in the intracellular fluid even though the extracellular fluid contains only a small concentration. This is true, for instance, for potassium ions.
Conversely, it is important to keep the concentrations of other ions very low inside the cell even though their concentrations in the extracellular fluid are great. This is especially true for sodium ions. Different substances that are actively transported through at least some cell membranes include sodium ions, potassium ions, calcium ions, iron ions, hydrogen ions, chloride ions, iodide ions, urate ions, several different sugars, and most of the amino acids.
Primary Active Transport Sodium-Potassium Pump
Sodium-potassium (Na+-K+) pump, a transport process that pumps
sodium ions outward through the cell membrane of all cells and at the same time pumps potassium ions from the outside to the inside. This pump is responsible for maintaining the sodium and potassium concentration differences across the cell membrane, as well as for
establishing a negative electrical voltage inside the cells.
The carrier protein is a complex of two separate globular proteins: a larger one called the (a) subunit, with a molecular weight of about 100,000, and a smaller one called the( b) subunit, with a molecular weight of about 55,000. Although the function of the smaller protein is not known (except that it might anchor the protein complex in the lipid membrane), the larger protein has three specific features that are important for the functioning of the pump:
1. It has three receptor sites for binding sodium ions on the portion of the protein that protrudes to the inside of the cell.
2. It has two receptor sites for potassium ions on the outside.
3. The inside portion of this protein near the sodium binding sites has ATPase activity.
To put the pump into perspective: When two potassium ions bind on the outside of the carrier protein and three sodium ions bind on the inside, the ATPase function of the protein becomes activated. This then cleaves
one molecule of ATP, splitting it to adenosine diphosphate (ADP) and liberating a high-energy phosphate bond of energy. This liberated energy is then believed to cause a chemical and conformational change in the protein carrier molecule, extruding the three sodium ions to the outside and the two potassium ions to the inside. As with other enzymes, the Na+-K+ ATPase pump can run in reverse.

Importance of the Na+-K+ Pump for Controlling Cell Volume.
One of the most important functions of the Na+-K+ pump is to control the volume of each cell. Without function of this pump, most cells of the body would swell until they burst. The mechanism for controlling the volume is as follows:Inside the cell are large numbers of proteins and other organic molecules that cannot escape from the cell. Most of these are negatively charged and therefore attract large numbers of potassium, sodium, and other positive ions as well. All these molecules and ions then cause osmosis of water to the interior of the cell. Unless this is checked, the cell will swell indefinitely until it bursts. The normal mechanism for preventing this is the Na+-K+ pump. Note again that this device pumps three Na+ ions to the outside of the cell for every two K+ ions pumped to the interior.
Electrogenic Nature of the Na+-K+ Pump.
The fact that the Na+-K+ pump moves three Na+ ions to the exterior for
every two K+ ions to the interior means that a net of one positive charge is moved from the interior of the cell to the exterior for each cycle of the pump.This creates positivity outside the cell but leaves a deficit of positive ions inside the cell; that is, it causes negativity on the inside. Primary Active Transport of Calcium Ions
Calcium ions are normally maintained at extremely low concentration in the intracellular cytosol of virtually all cells in the body, at a concentration about 10,000 times less than that in the extracellular fluid. This is achieved mainly by two primary active transport calcium pumps. One is in the cell membrane and pumps calcium to the outside of the cell. The other pumps calcium ions into one or more of the intracellular vesicular organelles of the cell, such as the sarcoplasmic reticulum of muscle cells and the mitochondria in all cells. In each of these instances, the carrier protein penetrates the membrane and functions as an enzyme ATPase, having the same capability to cleave ATP as the ATPase of the sodium carrier protein. The difference is that this protein has a highly specific binding site for calcium instead of for sodium.
Primary Active Transport of Hydrogen Ions At two places in the body, primary active transport of hydrogen ions is very important: in the gastric glands of the stomach and in the late distal tubules and cortical collecting ducts of the kidneys.
Secondary Active Transport— Co-Transport and Counter Transport
When sodium ions are transported out of cells by primary active transport, a large concentration gradient of sodium ions across the cell membrane usually develops—high concentration outside the cell and
very low concentration inside. This gradient represents a storehouse of energy because the excess sodium outside the cell membrane is always attempting to diffuse to the interior. Under appropriate conditions, this diffusion energy of sodium can pull other substances along with the sodium through the cell membrane. This phenomenon is called co-transport; it is one form of secondary active transport. For sodium to pull another substance along with it, a coupling mechanism is required. This is achieved by means of still another carrier protein in the cell membrane.
The carrier in this instance serves as an attachment point for both the sodium ion and the substance to be co-transported. Once they both are attached, the energy gradient of the sodium ion causes both the sodium ion and the other substance to be transported together to the interior of the cell.
In counter-transport, sodium ions again attempt to diffuse to the interior of the cell because of their large concentration gradient. However, this time, the substance to be transported is on the inside of the cell and must be transported to the outside. Therefore, the sodium ion binds to the carrier protein where it projects to the exterior surface of the membrane, while the substance to be counter-transported binds to the interior projection of the carrier protein. Once both have bound, a conformational change occurs, and energy released by the sodium ion moving to the interior causes the other substance to move to the exterior.
Co-Transport of Glucose and Amino Acids Along with Sodium Ions Glucose and many amino acids are transported into most cells against large concentration gradients; the mechanism of this is entirely by co-
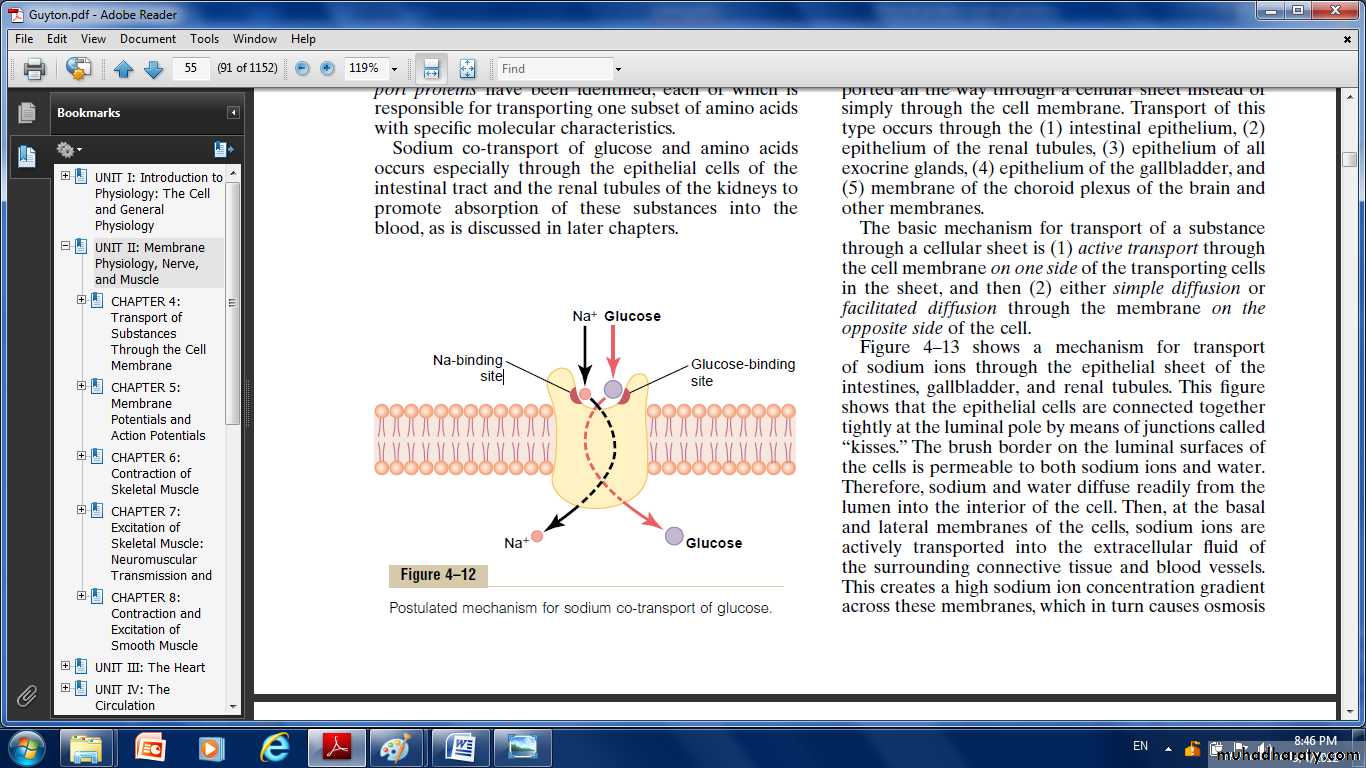
Fig. Postulated mechanism for sodium co-transport of glucose
transport, the transport carrier protein has two binding sites on its exterior side, one for sodium and one for glucose. Also, the concentration of sodium ions is very high on the outside and very low inside, which provides energy for the transport. A special property of the transport protein is that a conformational change to allow sodium movement to the
interior will not occur until a glucose molecule also attaches. When they both become attached, the conformational change takes place, and the sodium and glucose are transported to the inside of the cell at the same time. Hence, this is a sodium-glucose co-transport mechanism.
Sodium co-transport of the amino acids occurs in the same manner as for glucose, except that it uses a different set of transport proteins.
Other important co-transport mechanisms include co-transport of chloride ions, iodine ions, iron ions, and urate ions.
Sodium Counter-Transport of Calcium and Hydrogen Ions
Sodium-calcium counter-transport occurs through all or almost all cell membranes, with sodium ions moving to the interior and calcium ions to the exterior.
Sodium-hydrogen counter-transport occurs in the proximal tubules of the kidneys.
Active Transport Through Cellular Sheets: At many places in the body, substances must be transported all the way through a cellular sheet instead of simply through the cell membrane. Transport of this type occurs through the:
Intestinal epithelium.
Epithelium of the renal tubules.
Epithelium of the gallbladder.
Membrane of the choroid plexus of the brain and other membranes.
The basic mechanism for transport of a substance through a cellular sheet is:
(1) Active transport through the cell membrane on one side of the transporting cells in the sheet.
(2) Simple diffusion or facilitated diffusion through the membrane on the opposite side of the cell.
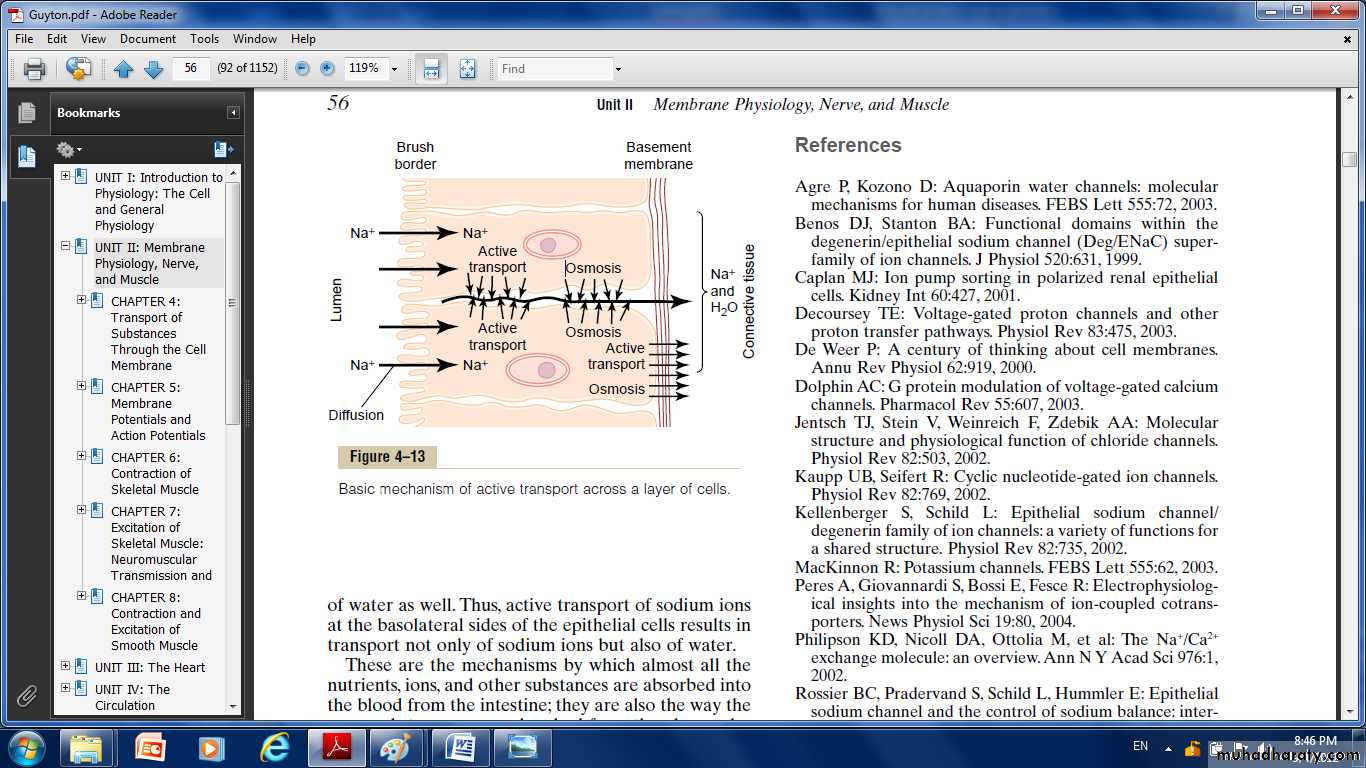
The epithelial cells are connected together tightly at the luminal pole by means of junctions called “kisses.” The brush border on the luminal surfaces of the cells is permeable to both sodium ions and water. Therefore, sodium and water diffuse readily from the lumen into the interior of the cell. Then, at the basal and lateral membranes of the cells, sodium ions are actively transported into the extracellular fluid of the surrounding connective tissue and blood vessels. This creates a high sodium ion concentration gradient across these membranes, which in turn causes osmosis of water as well. Thus, active transport of sodium ions at the basolateral sides of the epithelial cells results in transport not only of sodium ions but also of water. These are the mechanisms by which almost all the nutrients, ions, and other substances are absorbed into the blood from the intestine; they are also the way by which the same substances are reabsorbed from the glomerular filtrate by the renal tubules.