
hormone systems and their most important actions.
source of the sex hormones. Table 74–1 provides an overview of the different
endocrine tissues of the body, except for the placenta, which is an additional
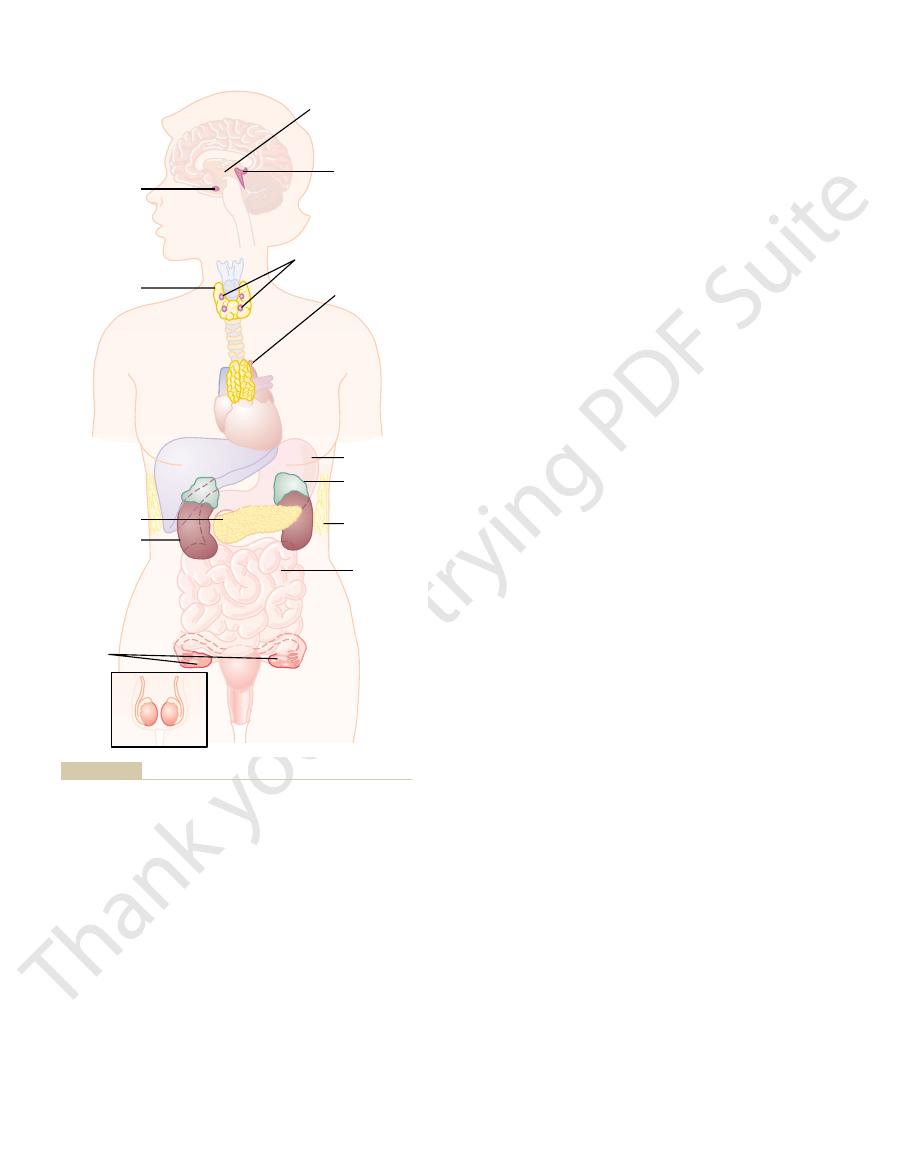
Figure 74–1 shows the anatomical loci of the major endocrine glands and
characteristics of the female body.
ovarian hormones
cortex, causing it to secrete adrenocortical hormones, and the
(ACTH)
have receptors for the hormone. For example,
target tissues,
tions in almost all the body’s cells.
thyroxine
(from the anterior pituitary gland) causes growth in most parts of the body, and
growth hormone
affect many different types of cells of the body; for example,
bind with receptors and initiate many reactions. Some endocrine hormones
throughout the body, including the nervous system in some cases, where they
The
of anterior pituitary hormones.
hypophysiotropic hormones,
(ADH), oxytocin,
eminence and secrete several neurohormones, including
thalamus, have axons that terminate in the posterior pituitary gland and median
in response to neural stimuli. The neuroendocrine cells, located in the hypo-
ger systems interact with one another to maintain homeostasis. For example,
hormone systems, keeping in mind that many of the body’s chemical messen-
In the next few chapters, we discuss mainly the endocrine and neuroendocrine
) produced by adipocytes are
34). Cytokine hormones (e.g.,
cytokines include the
function as autocrines, paracrines, or endocrine hormones. Examples of
receptors.
Autocrines
neighboring cells of a different type.
Paracrines
blood and influence the function of cells at another location in the body.
the body.
synaptic junctions and act locally to control nerve cell functions.
are released by axon terminals of neurons into the
The multiple activities of the cells, tissues, and
Coordination of Body
C
H
A
P
T
E
R
7
4
905
Introduction to Endocrinology
Functions by Chemical
Messengers
organs of the body are coordinated by the interplay
of several types of chemical messenger systems:
1. Neurotransmitters
2. Endocrine hormones are released by glands or specialized cells into the
circulating blood and influence the function of cells at another location in
3. Neuroendocrine hormones are secreted by neurons into the circulating
4.
are secreted by cells into the extracellular fluid and affect
5.
are secreted by cells into the extracellular fluid and affect the
function of the same cells that produced them by binding to cell surface
6. Cytokines are peptides secreted by cells into the extracellular fluid and can
interleukins and other lymphokines that are secreted
by helper cells and act on other cells of the immune system (see Chapter
leptin
sometimes called adipokines.
the adrenal medullae and the pituitary gland secrete their hormones primarily
antidiuretic hormone
and
which control the secretion
endocrine hormones are carried by the circulatory system to cells
(from the thyroid gland) increases the rate of many chemical reac-
Other hormones affect only specific
because only these tissues
adrenocorticotropic hormone
from the anterior pituitary gland specifically stimulates the adrenal
have specific effects on the female sex organs as well as on the secondary sexual
instances they are synthesized from cholesterol itself.
mones is similar to that of cholesterol, and in most
The chemical structure of steroid hor-
carried to their target tissues.
enter the circulatory system easily, where they are
peptide hormones are water soluble, allowing them to
kinases that initiate secretion of the hormone. The
receptor causes increased cyclic adenosine monophos-
instances, stimulation of an endocrine cell surface
depolarization of the plasma membrane. In other
increase in cytosolic calcium concentration caused by
In many cases, the stimulus for exocytosis is an
exocytosis.
until their secretion is needed. Secretion of the hor-
cytoplasm, and many are bound to the cell membrane
inactive fragments. The vesicles are stored within the
produce smaller, biologically active hormones and
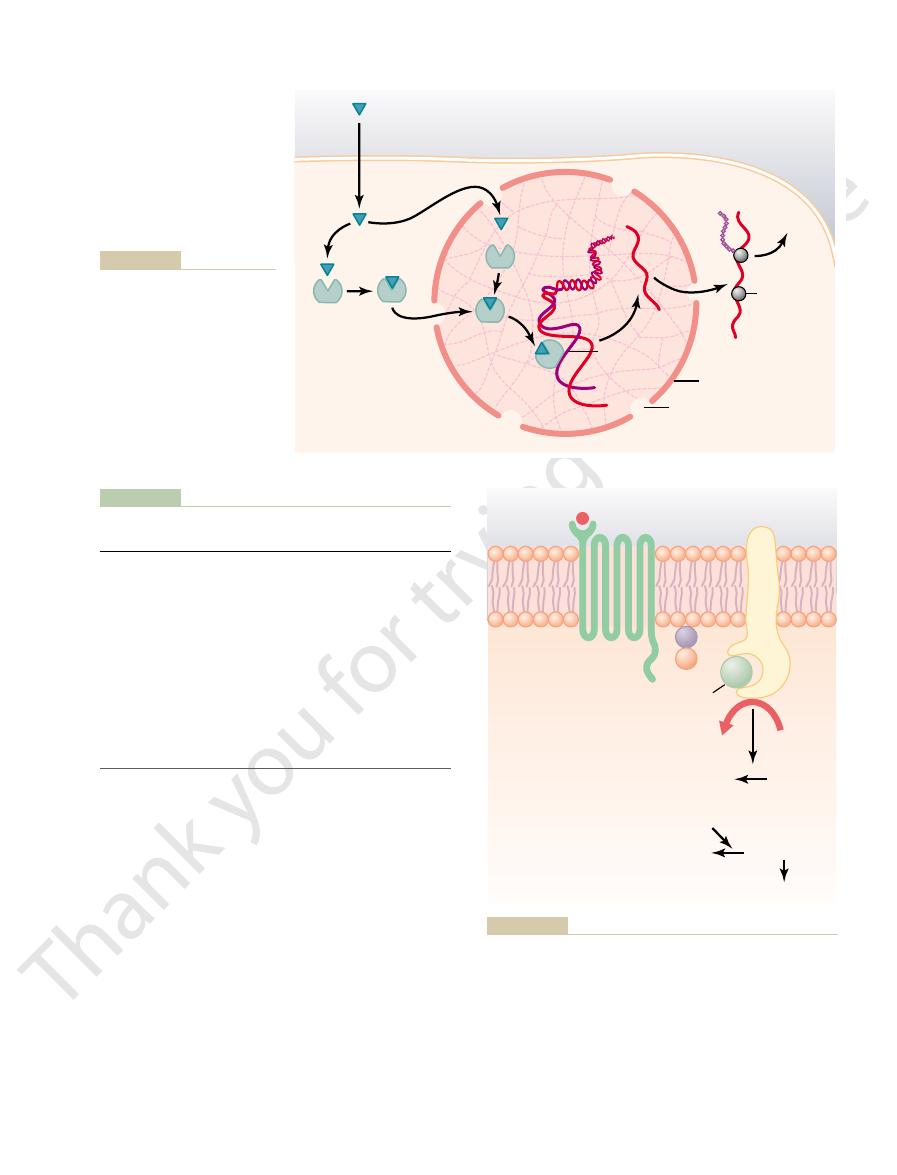
packaging into secretory vesicles. In this process,
These are then transferred to the Golgi apparatus for
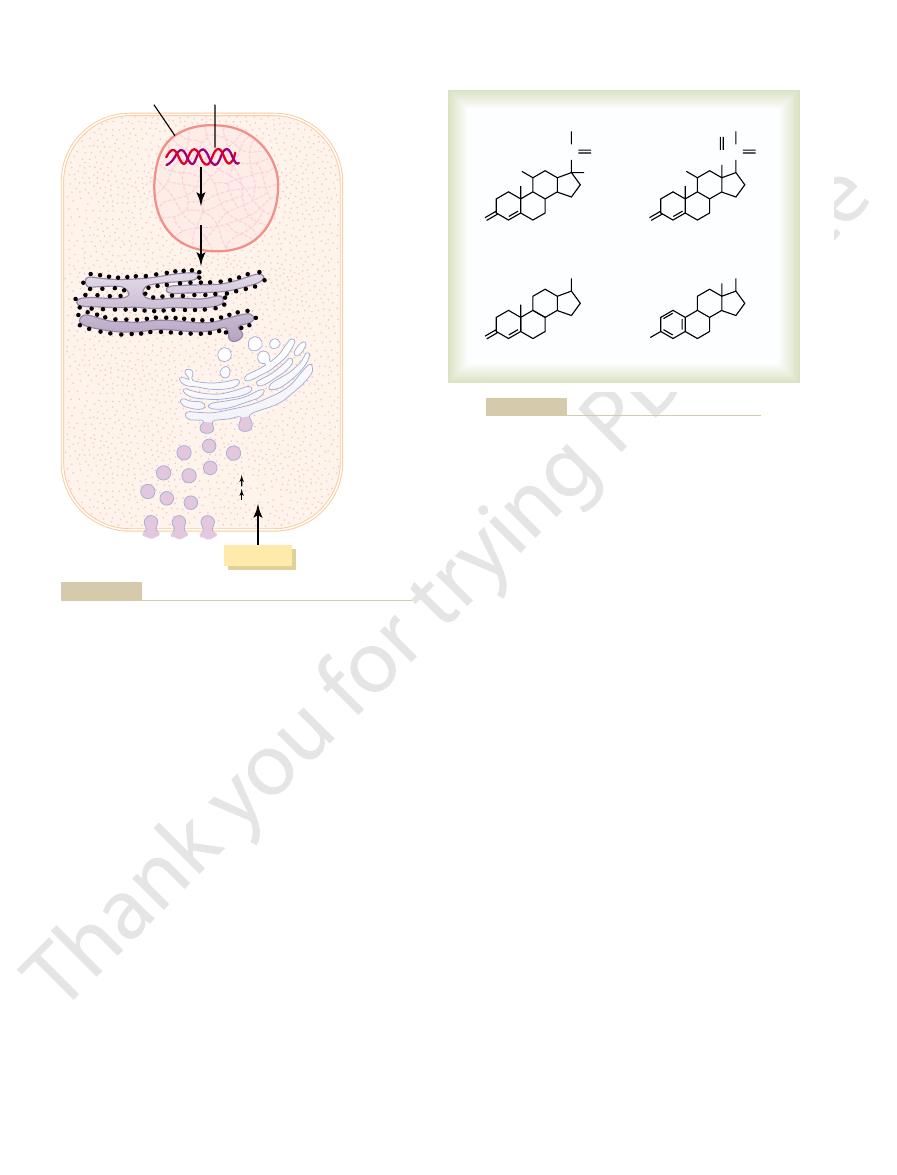
other proteins (Figure 74–2). They are usually synthe-
ferent endocrine cells, in the same fashion as most
than 100 amino acids are referred to as peptides.
amino acids are called proteins, and those with fewer
prolactin). In general, polypeptides with 100 or more
are polypeptides and proteins. These hormones range
Polypeptide and Protein Hormones Are Stored in Secretory Vesi-
polysaccharides or nucleic acid hormones.
norepinephrine). There are no known
Derivatives of the amino acid tyrosine,
progesterone), the testes (testosterone), and the
and aldosterone), the ovaries (estrogen and
many others (see Table 74–1).
parathyroid gland (parathyroid hormone), and
gland, the pancreas (insulin and glucagon), the
Proteins and polypeptides,
There are three general classes of hormones:
Synthesis of Hormones
Chemical Structure and
bohydrates for energy. And without the sex hormones,
creas, the body’s cells could use little of the food car-
become sluggish as well. Without insulin from the pan-
body would become sluggish, and the person would
thyroid gland, almost all the chemical reactions of the
Without thyroxine and triiodothyronine from the
without growth hormone, a person would be a dwarf.
balance, reproduction, and behavior. For instance,
olism, growth and development, water and electrolyte
regulating almost all body functions, including metab-
The multiple hormone systems play a key role in
906
Unit XIV
Endocrinology and Reproduction
sexual development and sexual functions would be
absent.
1.
including hormones
secreted by the anterior and posterior pituitary
2. Steroids secreted by the adrenal cortex (cortisol
placenta (estrogen and progesterone).
3.
secreted
by the thyroid (thyroxine and triiodothyronine)
and the adrenal medullae (epinephrine and
cles Until Needed.
Most of the hormones in the body
in size from small peptides with as few as 3 amino
acids (thyrotropin-releasing hormone) to proteins
with almost 200 amino acids (growth hormone and
Protein and peptide hormones are synthesized on
the rough end of the endoplasmic reticulum of the dif-
sized first as larger proteins that are not biologically
active (preprohormones) and are cleaved to form
smaller prohormones in the endoplasmic reticulum.
enzymes in the vesicles cleave the prohormones to
mones (as well as the inactive fragments) occurs when
the secretory vesicles fuse with the cell membrane and
the granular contents are extruded into the interstitial
fluid or directly into the blood stream by
phate (cAMP) and subsequently activation of protein
Steroid Hormones Are Usually Synthesized from Cholesterol and
Are Not Stored.
Pituitary gland
Adrenal
glands
Adipose
tissue
Thymus gland
Pineal gland
Hypothalamus
Parathyroid glands
(behind thyroid gland)
Stomach
Thyroid gland
Kidney
Pancreas
Ovaries
(female)
Small
intestine
Testes (male)
the body.
Figure 74–1
Anatomical loci of the principal endocrine glands and tissues of

Chapter 74
Introduction to Endocrinology
907
Table 74–1
Adipocytes
Leptin
Inhibits appetite, stimulates thermogenesis
Peptide
Cholecystokinin (CCK)
Stimulates gallbladder contraction and release of
Peptide
(Chapter 64)
bicarbonate and water
Small intestine
Secretin
Stimulates pancreatic acinar cells to release
Peptide
Stomach
Gastrin
Stimulates HCl secretion by parietal cells
Peptide
(Chapter 22)
blood pressure
Heart
Atrial natriuretic peptide (ANP)
Increases sodium excretion by kidneys, reduces
Peptide
Erythropoietin
Increases erythrocyte production
Peptide
1,25-Dihydroxycholecalciferol
Increases intestinal absorption of calcium and bone
Steroid
(Chapter 26)
angiotensin I (acts as an enzyme)
Kidney
Renin
Catalyzes conversion of angiotensinogen to
Peptide
Progesterone
See actions of progesterone from ovaries
Steroid
Estrogens
See actions of estrogens from ovaries
Steroid
fetal tissues as well as the mother’s breasts
Human somatomammotropin
Probably helps promote development of some
Peptide
(Chapter 82)
estrogens and progesterone by corpus luteum
Placenta
Human chorionic gonadotropin (HCG)
Promotes growth of corpus luteum and secretion of
Peptide
Progesterone
Stimulates secretion of “uterine milk” by the uterine
Steroid
(Chapter 81)
reproductive system, female breasts, and female
Ovaries
Estrogens
Promotes growth and development of female
Steroid
(Chapter 80)
system and male secondary sexual characteristics
Testes
Testosterone
Promotes development of male reproductive
Steroid
(Chapter 79)
increasing calcium absorption by the gut and
Parathyroid
Parathyroid hormone (PTH)
Controls serum calcium ion concentration by
Peptide
cells)
Increases synthesis and release of glucose from
Peptide
(Chapter 78)
this way controls carbohydrate metabolism
cells)
Promotes glucose entry in many cells, and in
Peptide
Pancreas
Insulin (
Adrenal medulla
Norepinephrine, epinephrine
Same effects as sympathetic stimulation
Amine
secretion, and hydrogen ion secretion
Aldosterone
Increases renal sodium reabsorption, potassium
Steroid
(Chapter 77)
metabolism of proteins, carbohydrates, and fats;
Adrenal cortex
Cortisol
Has multiple metabolic functions for controlling
Steroid
Calcitonin
Promotes deposition of calcium in the bones and
Peptide
(Chapter 76)
cells, thus increasing body metabolic rate
)
Increases the rates of chemical reactions in most
Amine
Thyroid
Thyroxine (T
Oxytocin
Stimulates milk ejection from breasts and uterine
Peptide
(Chapter 75)
vasopressin)
causes vasoconstriction and increased blood
Posterior pituitary
Antidiuretic hormone (ADH) (also called
Increases water reabsorption by the kidneys and
Peptide
luteum, and estrogen and progesterone synthesis
testes; stimulates ovulation, formation of corpus
Luteinizing hormone (LH)
Stimulates testosterone synthesis in Leydig cells of
Peptide
Follicle-stimulating hormone (FSH)
Causes growth of follicles in the ovaries and sperm
Peptide
Prolactin
Promotes development of the female breasts and
Peptide
hormones (cortisol, androgens, and aldosterone)
Adrenocorticotropic hormone (ACTH)
Stimulates synthesis and secretion of adrenocortical
Peptide
Thyroid-stimulating hormone (TSH)
Stimulates synthesis and secretion of thyroid
Peptide
(Chapter 75)
of most cells and tissues
Anterior pituitary
Growth hormone
Stimulates protein synthesis and overall growth
Peptide
Dopamine or prolactin-inhibiting factor (PIF)
Inhibits release of prolactin
Amine
Gonadotropin-releasing hormone (GnRH)
Causes release of LH and FSH
Growth hormone inhibitory hormone (GHIH)
Inhibits release of growth hormone
Peptide
Growth hormone–releasing hormone (GHRH)
Causes release of growth hormone
Peptide
(Chapter 75)
Corticotropin-releasing hormone (CRH)
Causes release of ACTH
Peptide
Hypothalamus
Thyrotropin-releasing hormone (TRH)
Stimulates secretion of TSH and prolactin
Peptide
Gland/Tissue
Hormones
Major Functions
Structure
Endocrine Glands, Hormones, and Their Functions and Structure
Chemical
(somatostatin)
hormones (thyroxine and triiodothyronine)
secretion of milk
maturation in Sertoli cells of testes
in ovaries
pressure
contractions
4
) and triiodothyronine (T
3
decreases extracellular fluid calcium ion
concentration
also has anti-inflammatory effects
(Chapter 60)
b
Glucagon (
a
the liver into the body fluids
kidneys and releasing calcium from bones
secondary sexual characteristics
endometrial glands and promotes development of
secretory apparatus of breasts
mineralization
(Chapter 64)
pancreatic enzymes
(Chapter 71)
functions are incredibly small. Their concentrations in
The concentrations of hormones
monal Secretion Rates.
full effect. Thus, each of the different hormones has its
roxine or growth hormone, may require months for
minutes; the actions of other hormones, such as thy-
seconds after the gland is stimulated, and they may
norepinephrine and epinephrine, are secreted within
Some hormones, such as
and Clearance from the Blood
Hormone Secretion, Transport,
with other substances.
Once the catecholamines enter the circulation, they
released from adrenal medullary cells by exocytosis.
stored in secretory granules, catecholamines are also
stored until secreted. Similar to the protein hormones
times more epinephrine than norepinephrine. Cate-
adrenal medulla, which normally secretes about four
hormones to the target tissues.
roxine-binding globulin,
mones combine with plasma proteins, especially
After entering the blood, most of the thyroid hor-
the amines are split from thyroglobulin, and the free
the thyroid gland. Hormone secretion occurs when
of the glandular cells. The thyroid hormones are
actions of enzymes in the cytoplasmic compartments
the adrenal medullary hormones, are formed by the
of hormones derived from tyrosine, the thyroid and
The two groups
Amine Hormones Are Derived from Tyrosine.
are synthesized, they simply diffuse across the cell
Because the steroids are highly lipid soluble, once they
synthesis of cholesterol in steroid-producing cells.
comes from the plasma, but there is also de novo
ulus. Much of the cholesterol in steroid-producing cells
stores of cholesterol esters in cytoplasm vacuoles can
storage in steroid-producing endocrine cells, large
structure (Figure 74–3).
rings and one cyclopentyl ring combined into a single
They are lipid soluble and consist of three cyclohexyl
908
Unit XIV
Endocrinology and Reproduction
Although there is usually very little hormone
be rapidly mobilized for steroid synthesis after a stim-
membrane and enter the interstitial fluid and then the
blood.
synthesized and stored in the thyroid gland and incor-
porated into macromolecules of the protein thy-
roglobulin, which is stored in large follicles within
hormones are then released into the blood stream.
thy-
which slowly releases the
Epinephrine and norepinephrine are formed in the
cholamines are taken up into preformed vesicles and
can exist in the plasma in free form or in conjugation
Onset of Hormone Secretion After a Stimulus, and Duration of
Action of Different Hormones.
develop full action within another few seconds to
own characteristic onset and duration of action—each
tailored to perform its specific control function.
Concentrations of Hormones in the Circulating Blood, and Hor-
required to control most metabolic and endocrine
the blood range from as little as 1 picogram (which is
one millionth of one millionth of a gram) in each mil-
liliter of blood up to at most a few micrograms (a few
Nucleus
DNA
Transcription
Golgi
apparatus
Storage
Storage
Secretion
Secretion
Stimulus
Extracellular
fluid
Ca
++
cAMP
Secretory
vesicles
Endoplasmic
reticulum
Packaging
Packaging
Synthesis
Synthesis
Translation
Translation
mRNA
hormone secretion often involves changes in intracellular calcium
Synthesis and secretion of peptide hormones. The stimulus for
Figure 74–2
or changes in cyclic adenosine monophosphate (cAMP) in the
cell.
Testosterone
Estradiol
Cortisol
Aldosterone
O
HO
OH
C
O
CH
2
OH
O
HO
HC C
O
O
CH
2
OH
O
OH
HO
OH
Chemical structures of several steroid hormones.
Figure 74–3

counting procedure. Then, using the formula just cited,
time, the plasma concentration of the radioactive
which gives one the rate of disappearance. At the same
centration in the plasma becomes steady. At this time,
Then the radioactive hormone is infused at a constant
be measured is tagged with a radioactive substance.
is the following: A purified solution of the hormone to
The usual procedure for making this measurement
hormone in each milliliter of plasma. Then, the meta-
To calculate this clearance rate, one measures (1)
liters of plasma cleared of the hormone per minute.
This
metabolic clearance rate.
hormone secretion into the blood. The second is
of a hormone in the blood. One of these is the rate of
Two factors can increase or decrease the concentration
target receptors or lost from the circulation. Binding
proteins serve as reservoirs, replenishing the concen-
The relatively large amounts of hormones bound to
proteins.
to plasma proteins. However, protein-bound hor-
the plasma exist free in solution. For example, more
in the blood mainly bound to plasma proteins. Usually
in contrast, circulate
Steroid and thyroid hormones,
stitial fluid, and ultimately to target cells.
where they diffuse out of the capillaries, into the inter-
ported from their sites of synthesis to target tissues,
Water-soluble hormones
Transport of Hormones in the Blood
release.
stages of sleep. In many cases, these cyclical variations
diurnal (daily) cycle, and sleep. For example, the secre-
changes, various stages of development and aging, the
Cyclical Variations Occur in Hormone Release.
tually, LH reaches an appropriate concentration, and
gen, which in turn causes more secretion of LH. Even-
itary before ovulation. The secreted LH then acts on
tion of the hormone. One example of this is the surge
positive feedback
few instances,
In a
hormones.
levels, including gene transcription and translation
Feedback regulation of hormones can occur at all
enough to slow further secretion of the hormone.
the target tissue. Therefore, only when the target tissue
The controlled variable is often not the secretory
target tissue.
release. In other words, the hormone (or one of its
the hormone, conditions or products resulting from the
at the target tissue. After a stimulus causes release of
negative feedback mech-
far appear to be closely controlled. In most instances,
occur throughout the day, all hormones studied thus
iological systems.
milligrams per day. We shall see later in this chapter
extremely small, usually measured in micrograms or
millionths of a gram) per milliliter of blood. Similarly,
Chapter 74
Introduction to Endocrinology
909
the rates of secretion of the various hormones are
that highly specialized mechanisms are available in the
target tissues that allow even these minute quantities
of hormones to exert powerful control over the phys-
Feedback Control of
Hormone Secretion
Negative Feedback Prevents Overactivity of Hormone Systems.
Although the plasma concentrations of many hor-
mones fluctuate in response to various stimuli that
this control is exerted through
anisms that ensure a proper level of hormone activity
action of the hormone tend to suppress its further
products) has a negative feedback effect to prevent
oversecretion of the hormone or overactivity at the
rate of the hormone itself but the degree of activity of
activity rises to an appropriate level will feedback
signals to the endocrine gland become powerful
steps involved in the synthesis of hormones and steps
involved in processing hormones or releasing stored
Surges of Hormones Can Occur with Positive Feedback.
occurs when the bio-
logical action of the hormone causes additional secre-
of luteinizing hormone (LH) that occurs as a result of
the stimulatory effect of estrogen on the anterior pitu-
the ovaries to stimulate additional secretion of estro-
typical negative feedback control of hormone secre-
tion is then exerted.
Superim-
posed on the negative and positive feedback control
of hormone secretion are periodic variations in
hormone release that are influenced by seasonal
tion of growth hormone is markedly increased during
the early period of sleep but is reduced during the later
in hormone secretion are due to changes in activity
of neural pathways involved in controlling hormone
(peptides and cate-
cholamines) are dissolved in the plasma and trans-
less than 10 per cent of steroid or thyroid hormones in
than 99 per cent of the thyroxine in the blood is bound
mones cannot easily diffuse across the capillaries and
gain access to their target cells and are therefore bio-
logically inactive until they dissociate from plasma
tration of free hormones when they are bound to
of hormones to plasma proteins greatly slows their
clearance from the plasma.
“Clearance” of Hormones
from the Blood
the rate of removal of the hormone from the blood,
which is called the
is usually expressed in terms of the number of milli-
the rate of disappearance of the hormone from the
plasma per minute and (2) the concentration of the
bolic clearance rate is calculated by the following
formula:
Metabolic clearance rate
= Rate of disappearance
of hormone from the plasma/Concentration of
hormone in each milliliter of plasma
rate into the blood stream until the radioactive con-
the rate of disappearance of the radioactive hormone
from the plasma equals the rate at which it is infused,
hormone is measured using a standard radioactive
the metabolic clearance rate is calculated.

effects on the postsynaptic cells. Although a few
calcium ions, and so forth. The altered movement of
for sodium ions, others for potassium ions, others for
ion channel–linked receptors
closing a channel for one or more ions. Some of these
in the structure of the receptor, usually opening or
naptic membrane. This almost always causes a change
norepinephrine, combine with receptors in the postsy-
transmitter substances, such as acetylcholine and
Virtually all the neuro-
types of interactions.
explain this, let us give a few examples of the different
activated receptor initiates the hormonal effects. To
This alters the function of the receptor itself, and the
Almost without exception, a hormone affects its target
Intracellular Signaling After Hormone
ing effects of the hormone.
the hormone. When this occurs, the target tissue
protein-manufacturing machinery of the target cell, or
and intracellular signaling proteins; that is, the stimu-
target tissue’s responsiveness to the hormone.
each case, receptor down-regulation decreases the
ized, or (5) decreased production of the receptors. In
interact with cell membrane receptors, (4) destruction
cell, away from the site of action of hormones that
intracellular protein signaling molecules, (3) tempo-
receptor molecules, (2) inactivation of some of the
down-regulation
decrease. This
For instance, increased hormone concentration and
course of their function, and at other times they
from minute to minute. The receptor proteins them-
does not remain constant from day to day, or even
The number of receptors in a target cell usually
one or more of the chromosomes.
The receptors for the thyroid
In the cell nucleus.
in the cytoplasm.
The primary receptors for
protein, peptide, and catecholamine hormones.
The
In or on the surface of the cell membrane.
The locations for the different types of hormone
that contain its specific receptors.
of hormone that will act on a particular tissue. The
specific for a single hormone; this determines the type
100,000 receptors. Also, each receptor is usually highly
Hormonal receptors are large proteins, and each cell
ates a cascade of reactions in the cell, with each stage
hormone combines with its receptor, this usually initi-
located in the cytoplasm or the nucleus. When the
membrane, whereas other hormone receptors are
tors for the hormones do not respond. Receptors
at the target cell. Cells that lack recep-
The first step of a hormone’s action is to bind to spe-
of Hormones
mones may be as long as 1 to 6 days.
for example, ranges between 20 and 100 minutes,
days. The half-life of adrenal steroids in the circulation,
in the blood is less than a minute.
For example, the half-life of angiotensin II circulating
liver, thus remaining in the blood for only a short time.
They are usually degraded by enzymes in the blood
tors are usually recycled back to the cell membrane.
hormone is then metabolized in the cell, and the recep-
the cell membrane hormone-receptor complex; the
cells by enzymatic processes that cause endocytosis of
“cleared” into the bile.
mones when the liver is diseased, because these hor-
For instance, this occurs for several of the steroid hor-
tration of the hormone in the circulating body fluids.
the urine. For certain hormones, a decreased metabolic
liver into the bile, and (4) excretion by the kidneys into
tissues, (2) binding with the tissues, (3) excretion by the
ways, including (1) metabolic destruction by the
Hormones are “cleared” from the plasma in several
910
Unit XIV
Endocrinology and Reproduction
clearance rate may cause an excessively high concen-
mones are conjugated mainly in the liver and then
Hormones are sometimes degraded at their target
Most of the peptide hormones and catecholamines
are water soluble and circulate freely in the blood.
and tissues and rapidly excreted by the kidneys and
Hormones that are bound to plasma proteins are
cleared from the blood at much slower rates and may
remain in the circulation for several hours or even
whereas the half-life of the protein-bound thyroid hor-
Mechanisms of Action
Hormone Receptors and Their
Activation
cific receptors
for some hormones are located on the target cell
becoming more powerfully activated so that even
small concentrations of the hormone can have a large
effect.
that is to be stimulated usually has some 2000 to
target tissues that are affected by a hormone are those
receptors are generally the following:
1.
membrane receptors are specific mostly for the
2. In the cell cytoplasm.
the different steroid hormones are found mainly
3.
hormones are found in the nucleus and are
believed to be located in direct association with
The Number and Sensitivity of Hormone Receptors Are Regu-
lated.
selves are often inactivated or destroyed during the
are reactivated or new ones are manufactured by
the protein-manufacturing mechanism of the cell.
increased binding with its target cell receptors some-
times cause the number of active receptors to
of the receptors can
occur as a result of (1) inactivation of some of the
rary sequestration of the receptor to the inside of the
of the receptors by lysosomes after they are internal-
Some hormones cause up-regulation of receptors
lating hormone induces greater than normal formation
of receptor or intracellular signaling molecules by the
greater availability of the receptor for interaction with
becomes progressively more sensitive to the stimulat-
Receptor Activation
tissues by first forming a hormone-receptor complex.
Ion Channel–Linked Receptors.
open (or close) channels
these ions through the channels causes the subsequent
(Figure 74–5). Leptin is a hormone
activity, others rely on enzymes that are closely asso-
is activated (or occasionally inactivated). Although
tor, an enzyme immediately inside the cell membrane
alytic or enzyme-binding site on the inside. When the
seven-transmembrane G protein–coupled receptors.
through the membrane only once, in contrast to the
These
closely associated with enzymes that they activate.
when activated, function directly as enzymes or are
Some receptors,
target tissues of the body.
of intracellular enzymes. This complex system of cell
receptor to an inhibitory or stimulatory G protein, a
teins). Thus, depending on the coupling of a hormone
proteins), whereas others are
units to form an inactive, membrane-bound trimeric G
itself by converting its bound GTP to GDP; then the
The signaling event is rapidly terminated when the
phospholipase C,
lular signaling proteins; these proteins, in turn, alter
trimeric G protein to associate with the cytoplasmic
subunit. When the receptor is activated, it undergoes
, and
In their inactive state,
guanosine nucleotides.
The trimeric G proteins are named for their ability
activity of an enzyme in the cytoplasm of the cell.
cellular part of the receptor, a conformational change
units. When the ligand (hormone) binds to the extra-
, and
include three (i.e., trimeric) parts—the
trude into the cell cytoplasm (especially the cytoplas-
cell membrane. Some parts of the receptor that pro-
protein–coupled receptors, all of which have seven
(Figure 74–4). There are more than 1000 known G
of target proteins (e.g., enzymes or ion channels) by
receptors, as discussed next.
activation of ion channel receptors, most hormones
Chapter 74
Introduction to Endocrinology
911
hormones may exert some of their actions through
that open or close ions channels do this indirectly by
coupling with G protein–linked or enzyme-linked
G Protein–Linked Hormone Receptors.
Many hormones
activate receptors that indirectly regulate the activity
coupling with groups of cell membrane proteins called
heterotrimeric GTP-binding proteins (G proteins)
transmembrane segments that loop in and out of the
mic tail of the receptor) are coupled to G proteins that
a, b
g sub-
occurs in the receptor that activates the G proteins and
induces intracellular signals that either (1) open or
close cell membrane ion channels or (2) change the
to bind
the
a, b
g subunits of G proteins form a complex
that binds guanosine diphosphate (GDP) on the
a
a conformational change that causes the GDP-bound
part of the receptor and to exchange GDP for guano-
sine triphosphate (GTP). Displacement of GDP by
GTP causes the
a subunit to dissociate from the
trimeric complex and to associate with other intracel-
the activity of ion channels or intracellular enzymes
such as adenylyl cyclase or
which
alters cell function.
hormone is removed and the
a subunit inactivates
a subunit once again combines with the b and g sub-
protein.
Some hormones are coupled to inhibitory G pro-
teins (denoted G
i
coupled to stimulatory G proteins (denoted G
s
pro-
hormone can either increase or decrease the activity
membrane G proteins provides a vast array of poten-
tial cell responses to different hormones in the various
Enzyme-Linked Hormone Receptors.
enzyme-linked receptors are proteins that pass
Enzyme-linked receptors have their hormone-binding
site on the outside of the cell membrane and their cat-
hormone binds to the extracellular part of the recep-
many enzyme-linked receptors have intrinsic enzyme
ciated with the receptor to produce changes in cell
function.
One example of an enzyme-linked receptor is the
leptin receptor
GTP activated
Receptor
Hormone
Cytoplasm
Extracellular
fluid
target protein
(enzyme)
GTP
G-protein
(active)
a
b
g
a
b
g
b
g
a
GDP
G-protein
(inactive)
membrane-bound target proteins (enzymes) that initiate intracellular signals.
subunits of the G protein and to interact with
subunit (to which the GTP is bound) to dissociate from the
complex associates with the receptor and is activated, with an exchange of guanosine triphosphate (GTP) for guanosine diphospha
G protein
Mechanism of activation of a G protein–coupled receptor. When the hormone activates the receptor, the inactive
Figure 74–4
a, b, and g
te
(GDP). This causes the
a
b and g
single type of membrane receptor. The second mes-
cellular effects of the hormone. Thus, the only direct
membrane. The cAMP then causes subsequent intra-
We noted earlier that one of the means by which hor-
the receptor regulates.
are tissue specific. Thus, the responses of different
teins is present, and many of these regulatory proteins
regulate are different in the various tissues. An intra-
hormone receptors, but the genes that the receptors
or altered cellular functions.
hormone has entered the cell, newly formed proteins
Therefore, minutes, hours, or even days after the
mation of messenger RNA (mRNA) (Figure 74–6).
(promoter) sequence of the DNA called the
the cytoplasm or nucleus. The activated hormone-
Because these hormones are lipid soluble, they readily
inside the cell rather than in the cell membrane.
mones, and vitamin D, bind with protein receptors
steroid hormones, thyroid hormones, retinoid hor-
Several hormones, including adrenal and gonadal
messenger.
cAMP, serves in a similar manner as a second
For a few peptide hormones, such as atrial natri-
cause these effects.
instead, the cAMP serves as a second messenger to
ity, as discussed in greater detail later. cAMP is called
cyclase catalyzes the formation of cAMP, which has a
end that protrudes to the interior of the cell. This
with a special transmembrane receptor, which then
control of cell function, is for the hormone to bind
Another example, one widely used in hormonal
synthesis of new proteins.
a result of activation of these intracellular enzymes,
JAK2 also leads to activation of other intracellular
genes to initiate protein synthesis. Phosphorylation of
teins, which activates transcription by leptin target
transducer and activator of transcription (STAT)
JAK2 complex to mediate intracellular signaling. The
The activated JAK2 molecules then phosphorylate
tion of the intracellular associated JAK2 molecules.
conformation, enabling phosphorylation and activa-
tor exists as a dimer (i.e., in two parts), and binding of
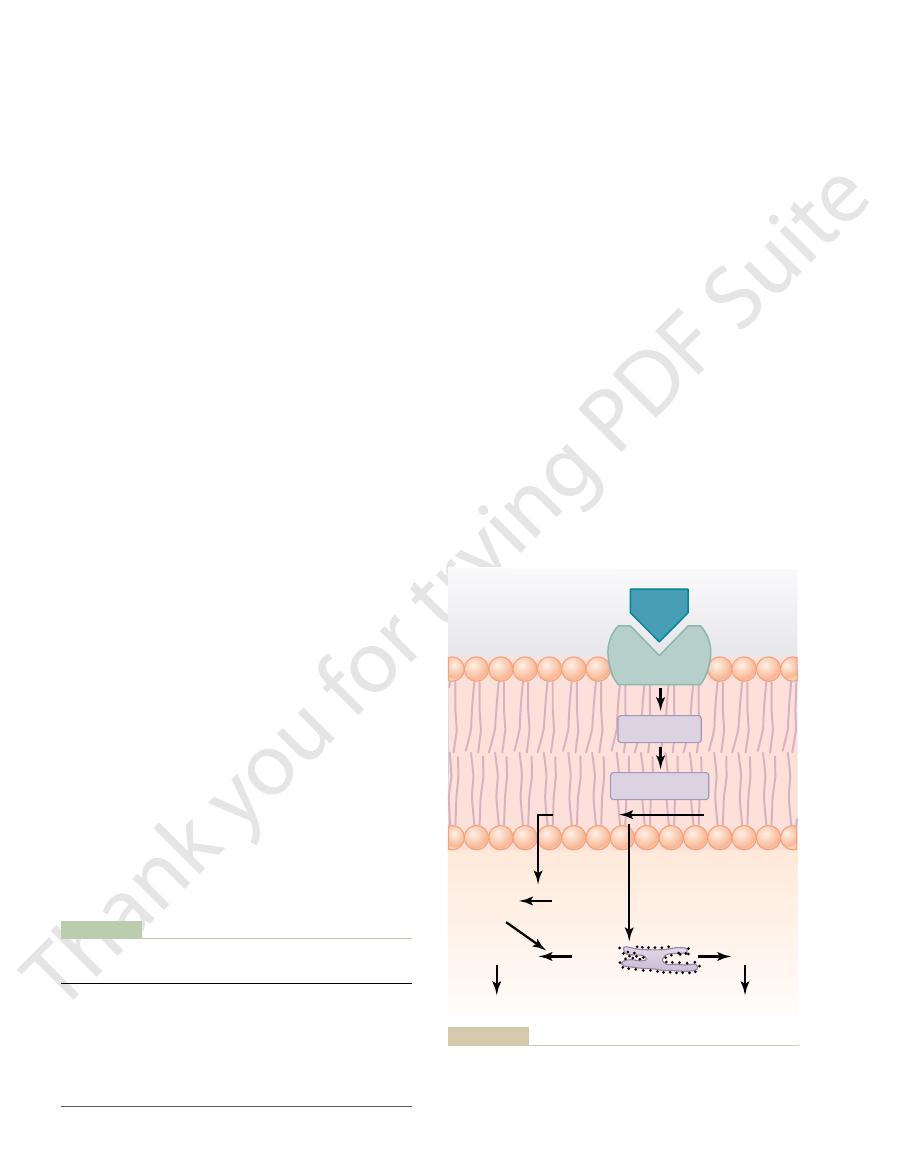
The leptin recep-
JAK2.
family,
janus kinase (JAK)
enzymes. In the case of the leptin receptor, one of the
71. The leptin receptor is a member of a large family
appetite and energy balance, as discussed in Chapter
effects, but it is especially important in regulating
912
Unit XIV
Endocrinology and Reproduction
secreted by fat cells and has many physiological
of cytokine receptors that do not themselves contain
enzymatic activity but signal through associated
signaling pathways occurs through a tyrosine kinase of
the
leptin to the extracellular part of the receptor alters its
other tyrosine residues within the leptin receptor–
intracellular signals include phosphorylation of signal
pro-
enzyme pathways such as mitogen-activated protein
kinases (MAPK) and phosphatidylinositol 3-kinase
(PI3K). Some of the effects of leptin occur rapidly as
whereas other actions occur more slowly and require
becomes the activated enzyme adenylyl cyclase at the
multitude of effects inside the cell to control cell activ-
a second messenger because it is not the hormone itself
that directly institutes the intracellular changes;
uretic peptide (ANP), cyclic guanosine monophos-
phate (cGMP), which is only slightly different from
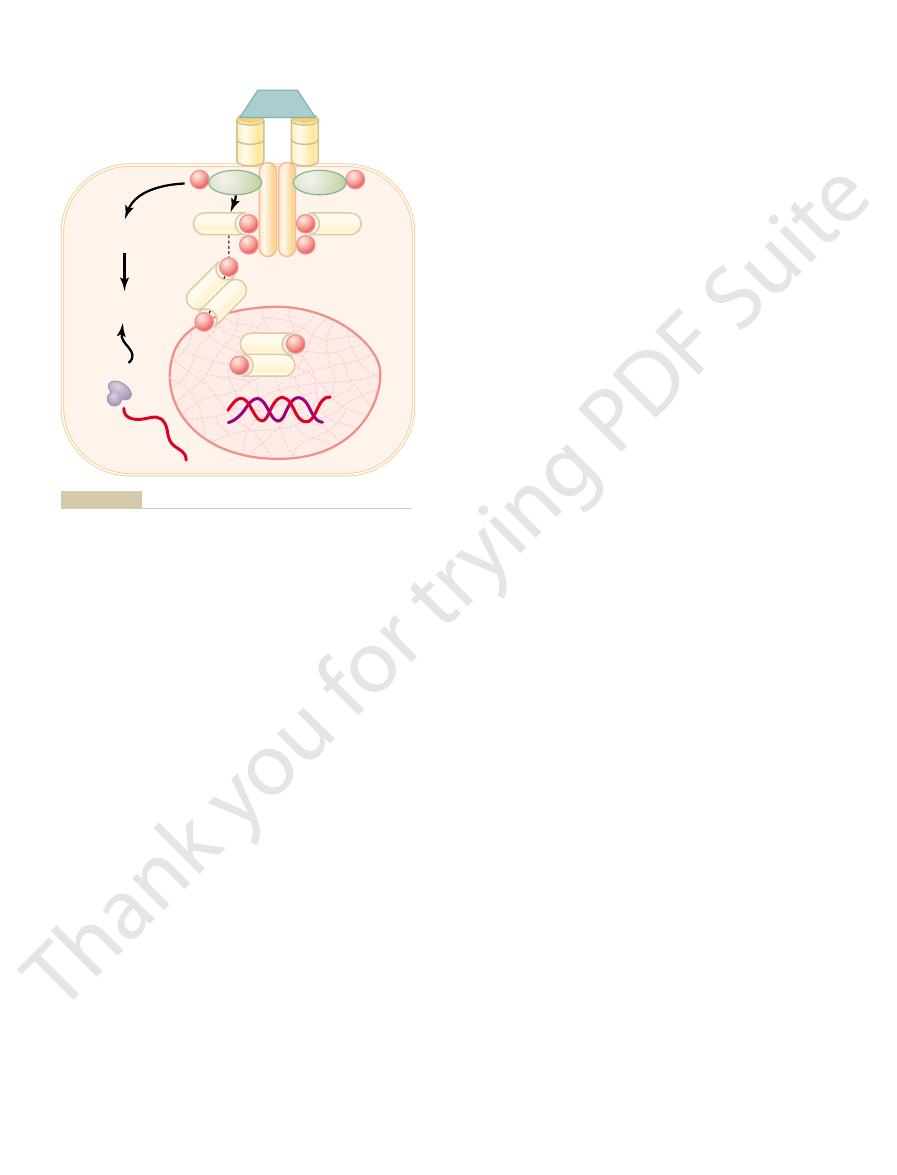
Intracellular Hormone Receptors and Activation of Genes.
cross the cell membrane and interact with receptors in
receptor complex then binds with a specific regulatory
hormone
response element, and in this manner either activates
or represses transcription of specific genes and for-
appear in the cell and become the controllers of new
Many different tissues have identical intracellular
cellular receptor can activate a gene response only if
the appropriate combination of gene regulatory pro-
tissues to a hormone are determined not only by the
specificity of the receptors but also by the genes that
Second Messenger Mechanisms
for Mediating Intracellular
Hormonal Functions
mones exert intracellular actions is to stimulate for-
mation of the second messenger cAMP inside the cell
effect that the hormone has on the cell is to activate a
senger does the rest.
Target gene
mRNA
Translation
Activation
of enzymes
Leptin receptor
Physiological
effects
Stat3
Stat3
P
P
Stat3
Jak2
Leptin
Jak2
Y
Y
Y
Y
Y
Y
P
P
P
P
P
P
Stat3
Stat3
Stat3
P
P
mediate some of the more rapid effects of leptin.
phorylation also activates several other enzyme systems that
tion of target genes and the synthesis of proteins. JAK2 phos-
transcription (STAT) proteins, which then activates the transcrip-
This causes phosphorylation of signal transducer and activator of
the extracellular part of the receptor, causing phosphorylation and
exists as a homodimer (two identical parts), and leptin binds to
An enzyme-linked receptor—the leptin receptor. The receptor
Figure 74–5
activation of the intracellular associated janus kinase 2 (JAK2).
lead to the cell’s response to the hormone.
cell, triggering biochemical reactions that ultimately
kinase,
cell. This then activates
(ATP) into cAMP inside the
the conversion of a small amount of cytoplasmic
brane-bound enzyme, by the G
G protein. Stimulation of adenylyl cyclase, a mem-
system, it is called a
G protein stimulates the adenylyl cyclase–cAMP
itself. Binding of the hormones with the receptor
lyl cyclase–cAMP second messenger system
their target tissues, and Figure 74–7 shows the adeny-
the adenylyl cyclase–cAMP mechanism to stimulate
Table 74–2 shows a few of the many hormones that use
different hormones. Two other especially important
Chapter 74
Introduction to Endocrinology
913
cAMP is not the only second messenger used by the
ones are (1) calcium ions and associated calmodulin
and (2) products of membrane phospholipid
breakdown.
Adenylyl Cyclase–cAMP Second
Messenger System
allows coupling of the receptor to a G protein. If the
G
s
protein, denoting a stimulatory
s
protein then catalyzes
adenosine triphosphate
cAMP-dependent protein
which phosphorylates specific proteins in the
Table 74–2
Some Hormones That Use the Adenylyl Cyclase–cAMP
Thyroid-stimulating hormone (TSH)
Follicle-stimulating hormone (FSH)
Adrenocorticotropic hormone (ACTH)
Second Messenger System
Angiotensin II (epithelial cells)
Calcitonin
Catecholamines (
b receptors)
Corticotropin-releasing hormone (CRH)
Glucagon
Human chorionic gonadotropin (HCG)
Luteinizing hormone (LH)
Parathyroid hormone (PTH)
Secretin
Somatostatin
receptor, epithelial cells)
Vasopressin (V
2
Target cell
Target cell
Diffusion
Diffusion
Lipophilic hormone
Lipophilic hormone
Extracellular fluid
Extracellular fluid
Proteins
Proteins
Ribosome
Ribosome
Nuclear envelope
Nuclear envelope
Nuclear pore
Nuclear pore
mRNA
mRNA
Hormone
response
element
Hormone
response
element
Hormone
receptor
complex
Hormone
receptor
complex
Cytoplasmic
receptor
Cytoplasmic
receptor
Nucleus
Nucleus
Nuclear
receptor
Nuclear
receptor
DNA
DNA
mRNA
mRNA
protein synthesis.
gene transcription, formation of
element (promoter) on the DNA.
binds to the hormone response
the hormone-receptor complex
hormone binds to the receptor in
tors in target cells. After the
steroids, with intracellular recep-
lipophilic hormones, such as
Figure 74–6
Mechanisms of interaction of
the cytoplasm or in the nucleus,
This either activates or inhibits
messenger RNA (mRNA), and
b
g
a
ATP
ATP
ADP Protein
Active
cAMP-
dependent
protein
kinase
Inactive
cAMP-
dependent
protein
kinase
cAMP
Adenylyl
cyclase
Protein – PO4
+
+
Cell’s response
Cytoplasm
Extracellular
fluid
Hormone
GTP
sine diphosphate; ATP, adenosine triphosphate.
many hormones exert their control of cell function. ADP, adeno-
Figure 74–7
Cyclic adenosine monophosphate (cAMP) mechanism by which
or inhibition of protein kinases. Activation of
multiple effects inside the cell, including activation
calcium, the calmodulin changes its shape and initiates
This protein has four calcium sites,
On entering a cell, calcium ions bind with the
calcium channels.
response to the entry of calcium into the cells. Calcium
throughout the body.
arachidonic acid,
effects, the lipid portion of DAG is
cell’s response (Figure 74–8). In addition to these
phorylates a large number of proteins, leading to the
DAG, the other lipid second messenger, activates
second messenger effects, such as smooth muscle con-
reticulum, and the calcium ions then have their own
The IP
diacylglycerol (DAG).
pholipids in the cell membrane, especially
This enzyme catalyzes the breakdown of some phos-
the inside projections of the receptors (Table 74–3).
increases their permeability to water.
mones. In epithelial cells of the renal tubules, cAMP
metabolic hormones thyroxine and triiodothyronine,
Thus, a thyroid cell stimulated by cAMP forms the
secretion by the cells, and altering cell permeability.
causing muscle contraction or relaxation, initiating
ating synthesis of specific intracellular chemicals,
tions are elicited in different target cells, such as initi-
cells have other enzymes. Therefore, different func-
ery—some cells have one set of enzymes, and other
The specific action that occurs in response to
G protein, a hormone can either increase or decrease
action in the cell. Thus, depending on the coupling of
adenylyl cyclase will be inhibited, reducing the forma-
so forth. In this way, even the slightest amount of
molecules of the third enzyme to be activated, and
enzyme to be activated, which can cause still more
adenylyl cyclase immediately inside the cell mem-
vates a third, and so forth. The importance of this
activated, which activates a second enzyme, which acti-
That is, first one enzyme is
cascade of enzymes.
Once cAMP is formed inside the cell, it usually acti-
914
Unit XIV
Endocrinology and Reproduction
vates a
mechanism is that only a few molecules of activated
brane can cause many more molecules of the next
hormone acting on the cell surface can initiate a pow-
erful cascading activating force for the entire cell.
If binding of the hormone to its receptors is coupled
to an inhibitory G protein (denoted G
i
protein),
tion of cAMP and ultimately leading to an inhibitory
the hormone receptor to an inhibitory or a stimulatory
the concentration of cAMP and phosphorylation of
key proteins inside the cell.
increases or decreases of cAMP in each type of target
cell depends on the nature of the intracellular machin-
whereas the same cAMP in an adrenocortical cell
causes secretion of the adrenocortical steroid hor-
The Cell Membrane Phospholipid Second
Messenger System
Some hormones activate transmembrane receptors
that activate the enzyme phospholipase C attached to
phos-
phatidylinositol biphosphate (PIP
2
), into two different
second messenger products: inositol triphosphate
(IP
3
) and
3
mobilizes
calcium ions from mitochondria and the endoplasmic
traction and changes in cell secretion.
the enzyme protein kinase C (PKC), which then phos-
which is the precursor for the prostaglandins and other
local hormones that cause multiple effects in tissues
Calcium-Calmodulin Second
Messenger System
Another second messenger system operates in
entry may be initiated by (1) changes in membrane
potential that open calcium channels or (2) a hormone
interacting with membrane receptors that open
protein calmodulin.
and when three or four of these sites have bound with
Some Hormones That Use the Phospholipase C Second
Table 74–3
Thyroid-releasing hormone (TRH)
Messenger System
Angiotensin II (vascular smooth muscle)
Catecholamines (
a receptors)
Gonadotropin-releasing hormone (GnRH)
Growth hormone–releasing hormone (GHRH)
Oxytocin
receptor, vascular smooth muscle)
Vasopressin (V
1
s response
Cell
Protein
Ca
Active
protein
kinase C
Inactive
protein
kinase C
Protein – PO
4
++
Endoplasmic reticulum
Cytoplasm
Receptor
Peptide
hormone
Cell membrane
Extracellular fluid
G protein
Phospholipase C
DAG
+
IP
3
PIP
2
Cell’
’s response
diacylglycerol; IP
which some hormones exert their control of cell function. DAG,
Figure 74–8
The cell membrane phospholipid second messenger system by
3
, inositol triphosphate; PIP
2
, phosphatidylinosi-
tol biphosphate.

hormone in the assayed fluid was small. Conversely, if
hormone to compete with the radioactive hormone,
radioactive hormone has bound with the antibody, it is
radioactive counting techniques. If a large amount of
remainder of the solution, and the quantity of radio-
Third, after binding has reached equilibrium, the
radioactive, that binds is proportional to its concen-
tity of each of the two hormones, the natural and the
of the antibody. In the process of competing, the quan-
and the hormone in the fluid to be assayed. Therefore,
must be met: There must be too little antibody to bind
radioactive isotope. However, one specific condition
Second, a small quantity of this antibody is (1)
follows. First, an antibody that is highly specific for the
The method of performing radioimmunoassay is as
radioimmunoassay.
ucts. This method is called
mones, their precursors, and their metabolic end prod-
sensitive method, however, was developed about 40
centrations by the usual chemical means.An extremely
Therefore, it was very difficult to measure these con-
one billionth of a milligram (1 picogram) per milliliter.
minute quantities; some concentrations are as low as
Measurement of Hormone
control functions for days or even weeks.
2. Once bound to the intranuclear receptors, the
metabolic activity in virtually all cells of the body.
proteins—probably 100 or more. Many of these
1. They activate the genetic mechanisms for the
Two important features of thyroid hormone func-
operators, as explained in Chapter 3.
located within the chromosomal complex, and they
nucleus. To accomplish this, these hormones first bind
thyroxine
The thyroid hormones
Transcription in the Cell Nucleus
Thyroid Hormones Increase Gene
vasopressin and norepinephrine.
peptide and amino acid–derived hormones, such as
several hours or even days. This is in marked contrast
Thus, the full action of the steroid hormone is charac-
the tubules and potassium secretion into the tubules.
about 45 minutes, proteins begin to appear in the renal
the sequence of events cited earlier ensues. After
aldosterone receptor protein. Therefore, in these cells,
plasm of renal tubular cells, which contain a specific
mones secreted by the adrenal cortex, enters the cyto-
aldosterone,
To give an example,
to form new proteins.
4. The mRNA diffuses into the cytoplasm, where it
mRNA.
DNA strands in the chromosomes, which activates
3. The combination binds at specific points on the
diffuses into or is transported into the nucleus.
2. The combined receptor protein–hormone then
membrane and enters the cytoplasm of the cell,
1. The steroid hormone diffuses across the cell
The sequence of events in steroid function is essen-
which in turn provide other functions of the cells.
enzymes, transport proteins, or structural proteins,
the target cells. These proteins then function as
ovaries, and testes—is to cause synthesis of proteins in
Another means by which hormones act—specifically,
Steroid Hormones Increase Protein Synthesis
Genetic Machinery of the Cell
Hormones That Act Mainly on the
both function and protein structure.
tal muscle contraction, as explained in Chapter 7. It is
tal muscle to activate troponin C, which causes skele-
actions of calmodulin. This is almost exactly the same
to activate the calmodulin system. But when the
mol/L, which is not enough
The normal calcium ion concentration in most cells
myosin kinase,
example, one specific function of calmodulin is to acti-
involved in the cell’s response to the hormone. For
phosphorylation, activation or inhibition of proteins
calmodulin-dependent protein kinases causes, via
Chapter 74
Introduction to Endocrinology
915
vate
which acts directly on the myosin
of smooth muscle to cause smooth muscle contraction.
of the body is 10
-8
to 10
-7
calcium ion concentration rises to 10
-6
to 10
-5
mol/L,
enough binding occurs to cause all the intracellular
amount of calcium ion change that is required in skele-
interesting that troponin C is similar to calmodulin in
the steroid hormones secreted by the adrenal cortex,
tially the following:
where it binds with a specific receptor protein.
the transcription process of specific genes to form
promotes the translation process at the ribosomes
one of the hor-
tubular cells and promote sodium reabsorption from
teristically delayed for at least 45 minutes—up to
to the almost instantaneous action of some of the
and triiodothyronine
cause increased transcription by specific genes in the
directly with receptor proteins in the nucleus itself;
these receptors are probably protein molecules
likely control the function of the genetic promoters or
tion in the nucleus are the following:
formation of many types of intracellular
are enzymes that promote enhanced intracellular
thyroid hormones can continue to express their
Concentrations in the Blood
Most hormones are present in the blood in extremely
years ago that revolutionized the measurement of hor-
Radioimmunoassay
hormone to be measured is produced.
mixed with a quantity of fluid from the animal con-
taining the hormone to be measured and (2) mixed
simultaneously with an appropriate amount of purified
standard hormone that has been tagged with a
completely both the radioactively tagged hormone
the natural hormone in the assay fluid and the radio-
active standard hormone compete for the binding sites
tration in the assay fluid.
antibody-hormone complex is separated from the
active hormone bound in this complex is measured by
clear that there was only a small amount of natural
and therefore the concentration of the natural
in the central nervous system. Ann N Y Acad Sci 1007:6,
Kelly MJ, Qiu J, Ronnekleiv OK: Estrogen modulation of G-
Biol 3:893, 2002.
tion and multivesicular-body sorting. Nat Rev Mol Cell
Katzmann DJ, Odorizzi G, Emr SD: Receptor downregula-
Ann N Y Acad Sci 994:111, 2003.
tion, and internalization of the ACTH receptor (MC2R).
Clark AJ, Baig AH, Noon L, et al: Expression, desensitiza-
expression. Physiol Rev 81:1269, 2001.
Aranda A, Pascual A: Nuclear hormone receptors and gene
the Cell. New York: Garland Science, 2002.
Alberts B, Johnson A, Lewis J, et al: Molecular Biology of
hormone levels.
mated using 96-well plates, and (3) it has proved to be
radioactive isotopes, (2) much of the assay can be auto-
The ELISA method has become widely used in clin-
antibody-hormone complexes. Therefore, the amount
assay methods, ELISA methods use excess antib-
detected. In contrast to competitive radioimmuno-
formation of many thousands of product molecules,
methods.
molecule. A third antibody (AB
or standards are added to each of the wells, followed
that is specific for the hormone being assayed. Samples
small wells. Each well is coated with an antibody (AB
74–10 shows the basic elements of this method, which
with the sensitivity of simple enzyme assays. Figure
mones. This test combines the specificity of antibodies
be used to measure almost any protein, including hor-
of hormone can often be assayed in this way.
fluid. As little as billionths or even trillionths of a gram
centration of the hormone in the “unknown” assayed
assay procedures with the standard curve, one can
radioactive counts recorded from the “unknown”
plotted, as shown in Figure 74–9. By comparing the
concentration levels. Then a “standard curve” is
“standard” solutions of untagged hormone at several
Fourth, to make the assay highly quantitative, the
natural hormone to compete for the binding sites.
bound, it is clear that there was a large amount of
916
Unit XIV
Endocrinology and Reproduction
only a small amount of radioactive hormone has
radioimmunoassay procedure is also performed for
determine within an error of 10 to 15 per cent the con-
Enzyme-Linked Immunosorbent
Assay (ELISA)
Enzyme-linked immunosorbent assays (ELISAs) can
is often performed on plastic plates that each have 96
1
)
by a second antibody (AB
2
) that is also specific for the
hormone but binds to a different site of the hormone
3
) is added that recog-
nizes AB
2
and is coupled to an enzyme that converts
a suitable substrate to a product that can be easily
detected by colorimetric or fluorescent optical
Because each molecule of enzyme catalyzes the
even very small amounts of hormone molecules can be
odies so that all hormone molecules are captured in
of hormone present in the sample or in the standard
is proportional to the amount of product formed.
ical laboratories because (1) it does not employ
a cost-effective and accurate method for assessing
References
protein-coupled receptor activation of potassium channels
2003.
2
4
8
16
32
64
100
90
80
70
60
50
40
30
20
10
0
128
Percent of antibody bound with
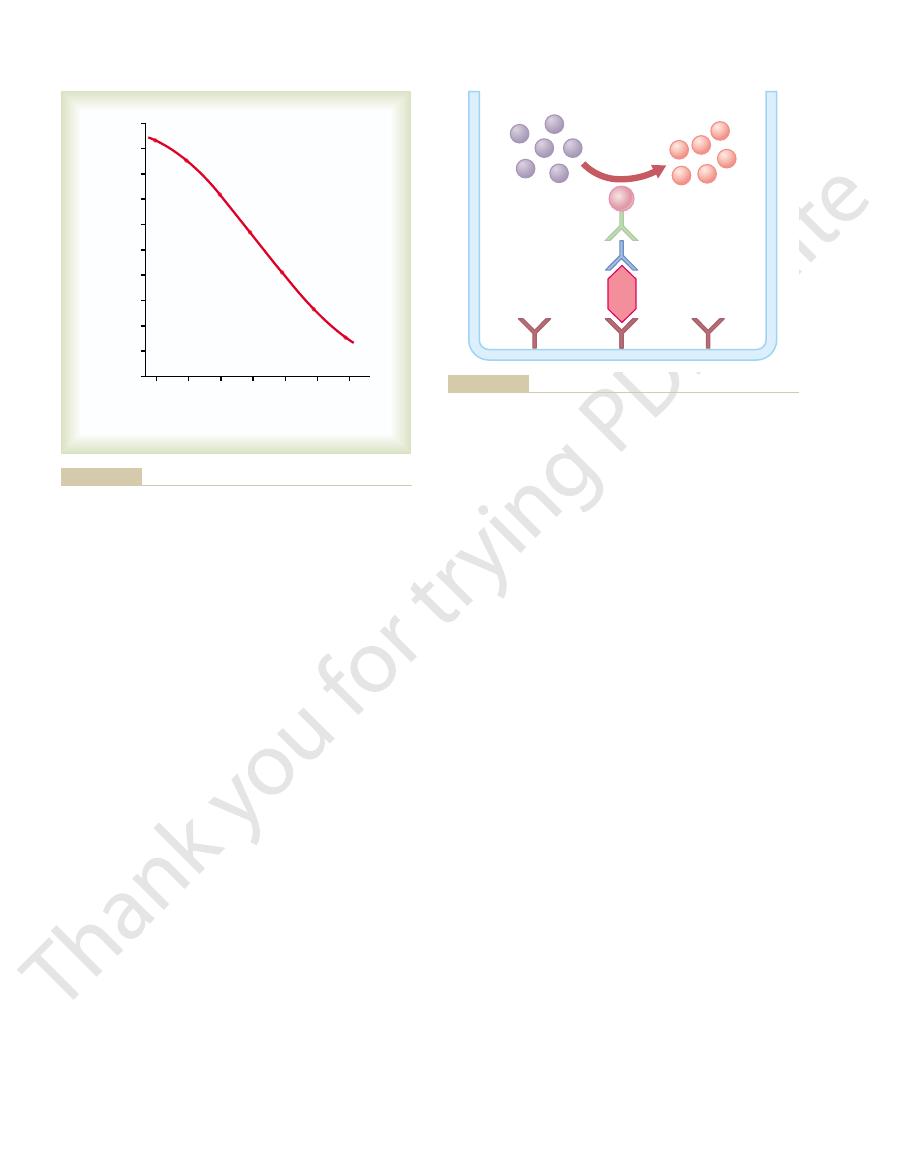
radioactive aldosterone
Aldosterone concentration in
test sample (ng/d)
Dr. Manis Smith.)
“Standard curve” for radioimmunoassay of aldosterone. (Courtesy
Figure 74–9
P
P
S
S
S
S
S
S
P
P
P
P
E
AB
3
AB
2
AB
1
H
the amount of hormone in the well if there are excess antibodies
product is measured using optical methods and is proportional to
fluorescent product (P) from a substrate (S). The amount of the
that catalyzes the formation of a colored
is an antibody that recognizes AB
are antibodies that recognize the hormone at different
(ELISA) for measuring the concentration of a hormone (H). AB
Figure 74–10
Basic principles of the enzyme-linked immunosorbent assay
1
and AB
2
binding sites, and AB
3
2
. E is an
enzyme linked to AB
3
in the well.

hormone action. Physiol Rev 81:1097, 2001.
Yen PM: Physiological and molecular basis of thyroid
molecular specificity. Physiol Rev 82:923, 2002.
hormone receptor interactions: physiological flexibility by
Vasudevan N, Ogawa S, Pfaff D: Estrogen and thyroid
84:137, 2004.
ated by distinct routes of protein kinase A. Physiol Rev
Tasken K, Aandahl EM: Localized effects of cAMP medi-
55:27, 2004.
proteins and G protein-coupled receptors. Annu Rev Med
Spiegel AM, Weinstein LS: Inherited diseases involving G
Physiol Rev 84:489, 2004.
model for convergence in cellular signaling pathways.
Spat A, Hunyady L: Control of aldosterone secretion: a
Physiol 66:315, 2004.
regulation by nuclear hormone receptors. Annu Rev
Privalsky ML: The role of corepressors in transcriptional
pathways in animal development. Nat Rev Genet 4:39,
Pires-daSilva A, Sommer RJ: The evolution of signaling
pathway. Nat Rev Drug Discov 3:555, 2004.
ity for immunosuppression: targeting the JAK/STAT
O’Shea JJ, Pesu M, Borie DC, Changelian PS: A new modal-
Physiol 283:R7, 2002.
leukocyte trafficking. Am J Physiol Regul Integr Comp
Olson TS, Ley K: Chemokines and chemokine receptors in
N Y Acad Sci 1024:213, 2004.
trafficking and gene targeting by steroid receptors. Ann
Nagaich AK, Rayasam GV, Martinez ED, et al: Subnuclear
16:251, 2001.
brane, cytosolic, and nuclear effects. News Physiol Sci
Nadal A, Diaz M, Valverde MA: The estrogen trinity: mem-
protein-linked signaling. Physiol Rev 79:1373, 1999.
Morris AJ, Malbon CC: Physiological regulation of G
Physiol Rev 83:965, 2003.
steroid action: controversies, questions, and answers.
Lösel RM, Falkenstein E, Feuring M, et al: Nongenomic
WB Saunders, 2003.
Williams Textbook of Endocrinolog, 10th ed. Philadelphia:
Larsen PR, Kronenberg HM, Melmed S, Polonsky KS:
guanylyl cyclase-A. Circ Res 93:700, 2003.
membrane guanylyl cyclase receptors, with a focus on
Kuhn M: Structure, regulation, and function of mammalian
Chapter 74
Introduction to Endocrinology
917
2003.