Physiology
Dr. Basim Mohamad Alwan Lecture (1)
INTRODUCTION
The nervous system is unique in the vast complexity of thought
processes and control actions it can perform. It receives each minute
literally millions of bits of information from the different sensory
nerves and sensory organs and then integrates all these to determine
responses to be made by the body. The nervous system contains more
than 100 billion neurons and consists of the central nervous system
(CNS) and the peripheral nerves (fig. 1-1).
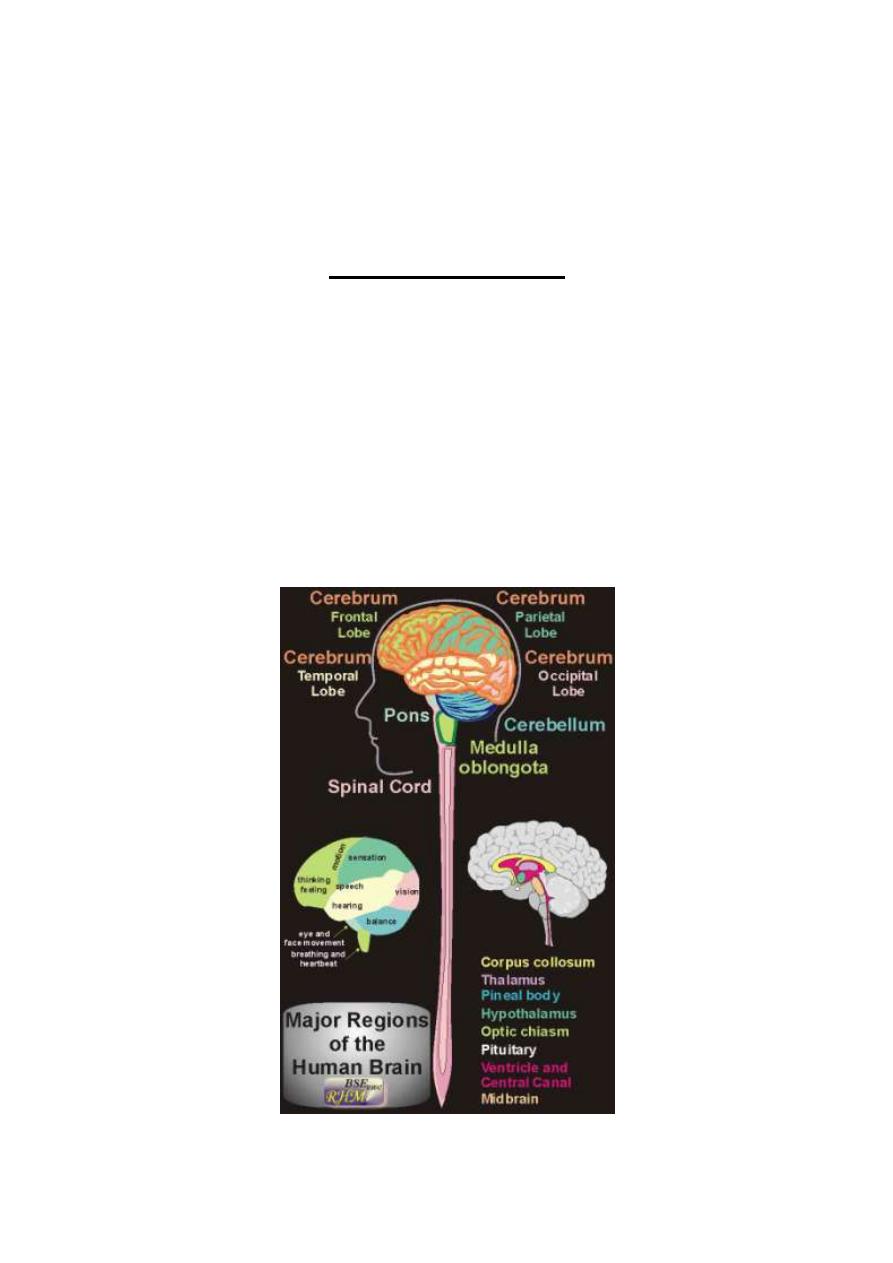
Figure 1-1: General view of the brain, spinal cord and spinal nerves

The central nervous system (the neural axis or neuraxis) consists of the
brain and the spinal cord.
Anatomically, the brain comprises the cerebrum which consists of two
cerebral hemispheres, the cerebellum, and the brain stem. The
brainstem consists of the midbrain, the pons and the medulla oblongata.
The CNS contains the nerve centers which receive and process the
nervous signals, then formulate the response to these signals. The
peripheral nerves are divided into cranial and spinal nerves. The cranial
nerves are twelve pairs of nerves, which arise from the brain and
emerge out through foramina in the bones of the cranium (skull). The
spinal nerves are 31 pairs of nerves that arise from the spinal cord and
emerge out through foramina in the vertebral column. Anatomically,
the spinal nerves are sorted out into 5 groups according to their site of
origin from the spinal cord which are 8 cervical pair, 12 dorsal pairs, 5
lumbar pairs, 5 sacral pairs and one cooccygeal pair.
DIVISIONS OF THE NERVOUS SYSTEM
The nervous system comprises three major systems;
I. THE AUTONOMIC NERVOUS SYSTEM
: Is the part of the nervous
system which is concerned with the involuntary control of the visceral
activity. It includes sympathetic, parasympathetic and enteric
divisions.
II. THE SOMATIC NERVOUS SYSTEM:
Is the part of the nervous
system which is concerned with conscious perception of different
sensations, and voluntary control of the muscular activity.

This system is divided into two divisions;
(i) SENSORY DIVISION:
Which is concerned with conscious
perception of somatic sensations? It includes the sensory (afferent)
nerves, the sensory (ascending) tracts inside the CNS, the sensory
reticular formation, the thalamus and the sensory cerebral cortex.
(ii) MOTOR DIVISION:
Which is concerned with voluntary control of
muscular activity? It includes the motor cerebral cortex, the basal
ganglia, the cerebellum, the motor reticular formation, the motor
(descending) tracts inside the CNS and the motor (efferent) nerves.
(iii). THE INTEGRATIVE NERVOUS SYSTEM: Is the part of the
nervous system which is concerned with the sophisticated functions of
the brain. These functions include memory, thinking, learning,
language, speech, emotions and general behavior. The main parts of the
integrative division are the cortical association areas and the limbic
system.
All these three systems and divisions are interconnected and their
functions are integrated together and with other systems in the body.
The basic functional unit in the nervous system is the reflex action.
A reflex action is an involuntary action in response to a stimulus e.g. a.
painful stimulus applied to the hand leads to reflex withdrawal of the
arm (the withdrawal reflex).
The basic structural unit of the nervous system which is capable of
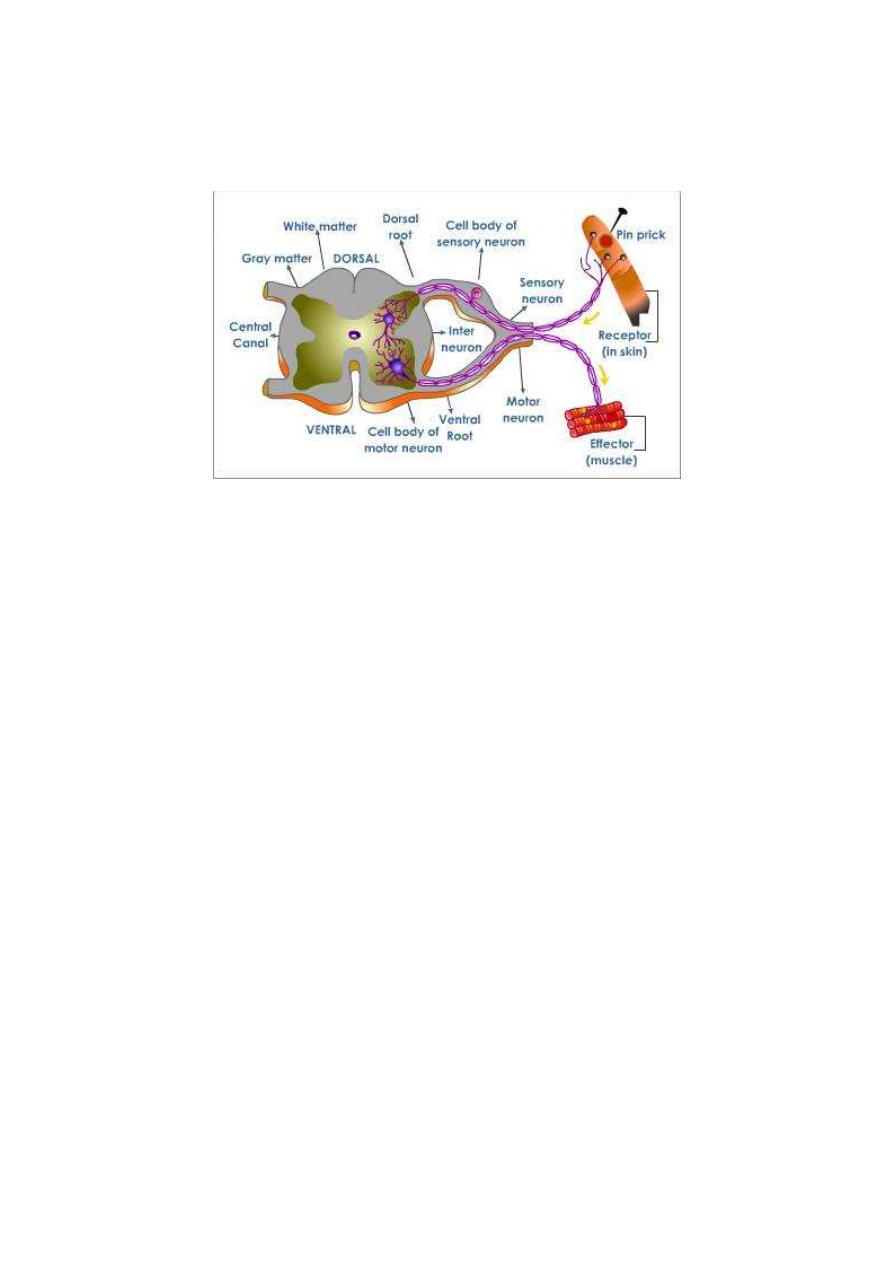
conducting a reflex action is the reflex arc (fig. 1-2). A reflex arc;
consists of 5 components:
Figure 1-2: The five basic components of a reflex arc
1. Receptor: A sensor which is excited by the stimulus.
2. Afferent nerve: Which conveys input signals to the CNS? The
afferent nerve is also called the sensory nerve,
3. Center: A collection of neurons that receive the sensory
information and issue the order for proper response.
4. Efferent nerve: A nerve that conveys output signals from the CNS
to the effector organ. The efferent nerve is either a motor nerve to a
muscle or a secretary nerve to a gland.
5. Effector organ: A muscular or glandular structure which receives
the final order and executes the reflex response.

NEURAL SYNAPSES
A synapse is the junctional area between a nerve terminal and another
cell. If the second cell is a neuron the synapse is then called a "neural
or neuronal synapse".
The axon of a neuron conducts impulses away from the cell body to
relay onto another cell at the synapses (fig. 1.3). The axon branches
extensively near its end, giving off 1000 branches on the average. Each
branch ends in a nerve terminal. This terminal is a disc-like expansion
called the synaptic knob (the terminal button or the end foot). There
is a gap between the nerve terminal and the adjacent neuron 3 0 - 50 nm
wide called the synaptic cleft. So, at the neural synapse there is
contiguity but no continuity of the two adjacent neurons.
The neuron which conducts impulses to the synapse is called the
"presynaptic neuron'' or "input neuron" and that which conducts
impulses away from the synapse is called the "postsynaptic neuron"
or "output neuron". The synaptic knobs of the presynaptic neuron
contain vesicles called synaptic or transmitter vesicles which contain
the chemical transmitter of the neuron. A polypeptide called synapsin
is found in the walls of the vesicles which bind the transmitter vesicles
to the cytoskeleton keeping them in the cytoplasm away from the release
sites on the presynaptic membrane.
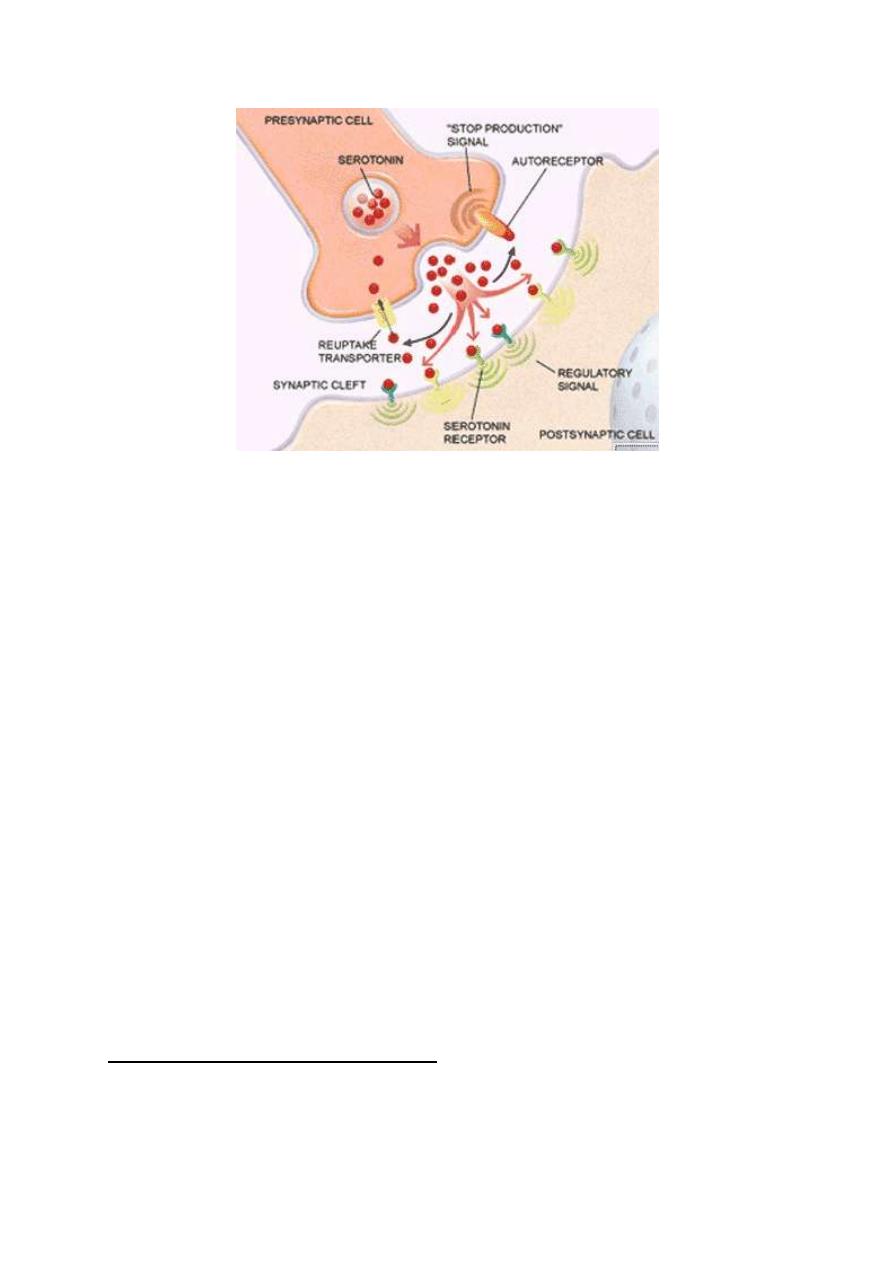
Figure 1-3: Neural synapse
THE IMPORTANCE OF SYNAPSES IN THE NERVOUS SYSTEM
Synapses act as "unidirectional valves" in the nervous pathways, i.e.
they allow the flow of impulses from the pre to the postsynaptic
neurons only. This ensures the flow of impulses in the nervous
pathways in the forward "orthodromic" direction only. Any impulse
that travels along a neuron in the opposite "antidromic" direction
cannot be transmitted to the next neuron because it dies off at the first
set of synapses it meets.
Also, synapses are the sites in the nervous pathways at which
transmission of impulses can be most easily influenced. At the synapse,
transmission of impulses can be accelerated, slowed down, or blocked
by physiological, pathological or pharmacological influences.
CLASSIFICATION OF SYNAPSES
Synapses could be classified according to either their location between
the pre and postsynaptic neurons (histological classification), or the
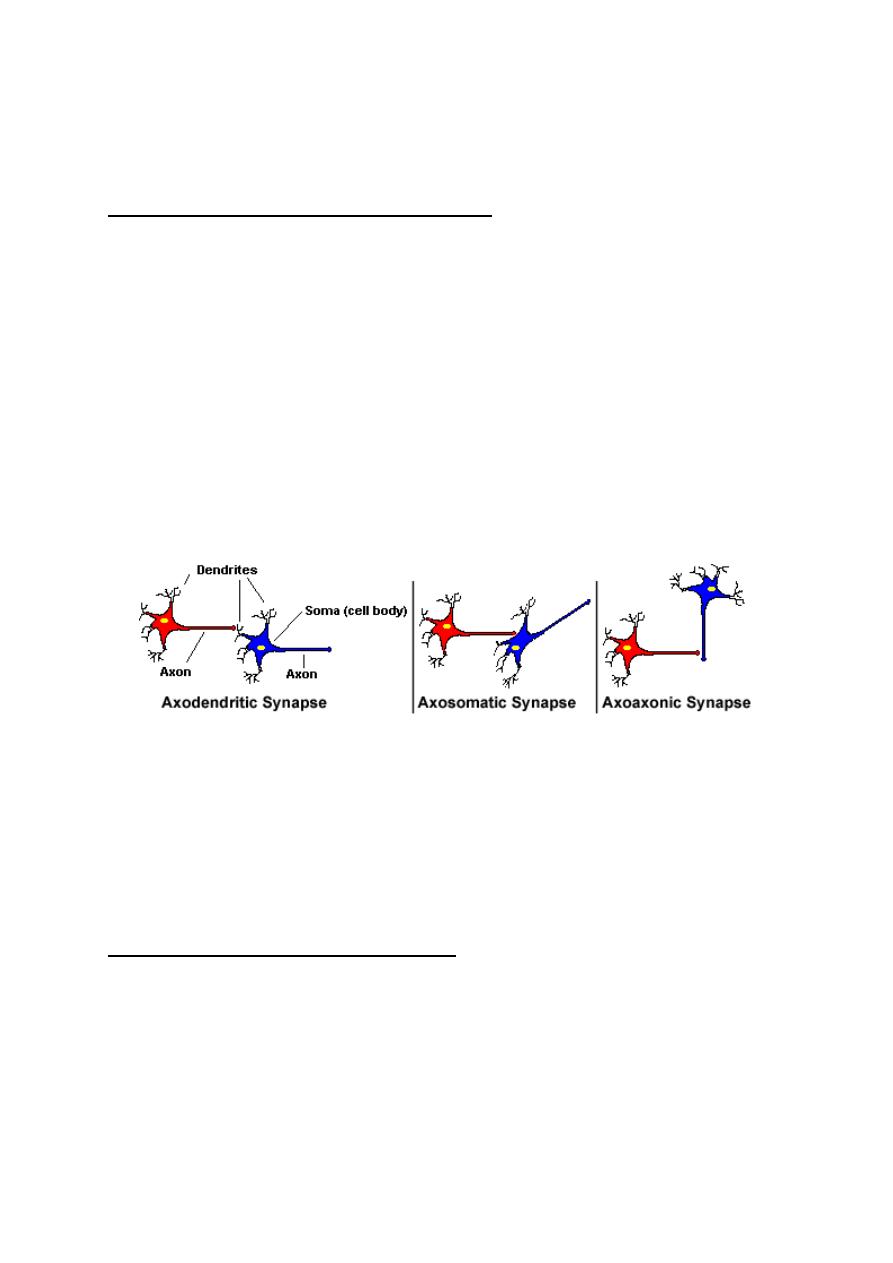
mechanism of transmission of impulses across them (physiological
classification),
HISTOLOGICAL CLASSIFICATION
According to this classification, synapses are classified into three types
(fig.1-4):
1. Axodendritic synapses: These are synapses between the axon
terminals of the presynaptic neuron and the dendrites of the
postsynaptic neuron.
2. Axosomatic synapses: These are synapses between the axon
terminals of the presynaptic neuron and the soma of the postsynaptic
neuron
Figure 1-4: histological classification of synapses
3. Axoaxonic synapses: These are synapses between the axon
terminals of the presynaptic neuron and the axon of the postsynaptic
neuron.
PHYSIOLOGICAL CLASSIFICATION
According to this classification, synapses are classified into three
types:
1. CHEMICAL SYNAPSES:
In these synapses, transmission of signals
occurs by releasing a ''chemical transmitter" from the presynaptic

terminal into the synaptic cleft. The transmitter then acts on specific
receptors on the postsynaptic membrane to generate postsynaptic
potential. There are more than 40 different synaptic transmitters in the
CNS which are either small molecule rapidly acting (acetylcholine) or
large molecule slowly acting (substance P).
Chemical synapses are the only type of synapses found in the
human nervous system.
2. ELECTRICAL SYNAPSES:
In these synapses, there are gap
junctions between the pre and postsynaptic membranes which allow
the transmission of the depolarization wave directly from the pre to the
postsynaptic membrane.
3.
CONJOINT SYNAPSES (ELECTROCHEMICAL):
In these synapses,
transmission of impulses occurs by both mechanisms electrical and
chemical. They are found in some fish and invertebrates.
THE MECHANISM OF RELEASE OF TRANSMITTER AT THE
CHEMICAL SYNAPSES
When the action potential reaches the nerve terminal, it opens the calcium
gates allowing Ca
2+
influx from the extracellular fluid into the cytoplasm.
Ca
2+
induces the phosphorylation of synapsin. This detaches the
synaptic vesicles from their binding to the cytoskeleton. The vesicles get
attached and fused to specific release sites on the presynaptic membrane.
The release sites then rupture and the chemical transmitter is released
into the synaptic cleft. This process is a passive process.
THE MECHANISM OF ACTION OF THE CHEMICAL TRANSMITTER
The transmitter moves in the fluid in the synaptic cleft by simple

diffusion to the receptors on the postsynaptic membrane. It activates these
receptors to generate postsynaptic potential (PSP). There are two types
of PSPs; excitatory and inhibitory:
1. THE EXCITATORY POSTSYNAPTIC POTENTIALS (EPSPs)
EPSPs are produced by depolarization of the postsynaptic membrane
due to sodium influx. The action of the chemical transmitter is to open
the gated sodium channels leading to sodium influx. Some transmitters
produce EPSPs by closing K
+
channels (membrane depolarization).
2. THE INHIBITORY POSTSYNAPTIC POTENTIALS (IPSPs
)
IPSPs are produced by hyperpolarization of the postsynaptic membrane
due to C1
-
influx and / or K
+
efflux. The action of the transmitter in this
case is to open the gated chloride and / or potassium channels. The IPSPs
decrease the excitability of the postsynaptic neuron and resist the
development of any action potential in it.
The excitatory and inhibitory postsynaptic potentials do not obey the
all or none rule. They can be summated either temporally or spatially.
SUMMATION OF THE POSTSYNAPTIC POTENTIALS
There are two ways of summation of the postsynaptic potentials;
temporal and spatial summation.
1. TEMPORAL SUMMATION
This is summation of the postsynaptic potentials produced by a train of
impulses on one presynaptic terminal, reaching the same synapse one
shortly after the other (fig. 1-5). Each time an impulse reaches the
synapse it creates a PSP which is summated with other PSPs in its
magnitude.

Figure 1-5: The mechanism of temporal summation.
Temporal summation is possible because the opening of a ligand-gated
channel lasts for about one ms (millisecond) whilst the PSP produced
by this opening lasts for about 15 ms. In this way any other opening of
the same channel within 15 ms would produce another PSP that will
potentiate the previous one.
2. SPATIAL SUMMATION
This is summation of postsynaptic potentials produced by multiple impulses
in several presynaptic terminals which reach several synapses at the same
moment (fig. 1-6). A large number of postsynaptic spots are stimulated
simultaneously. This increases the stimulated surface area of the
postsynaptic neuron. The simultaneously generated postsynaptic potentials
potentiate each other. In most cases in vivo, both types of summation
occur at the same time (temporospatial summation) where multiple
impulses arrive one shortly after another at several presynaptic terminals.
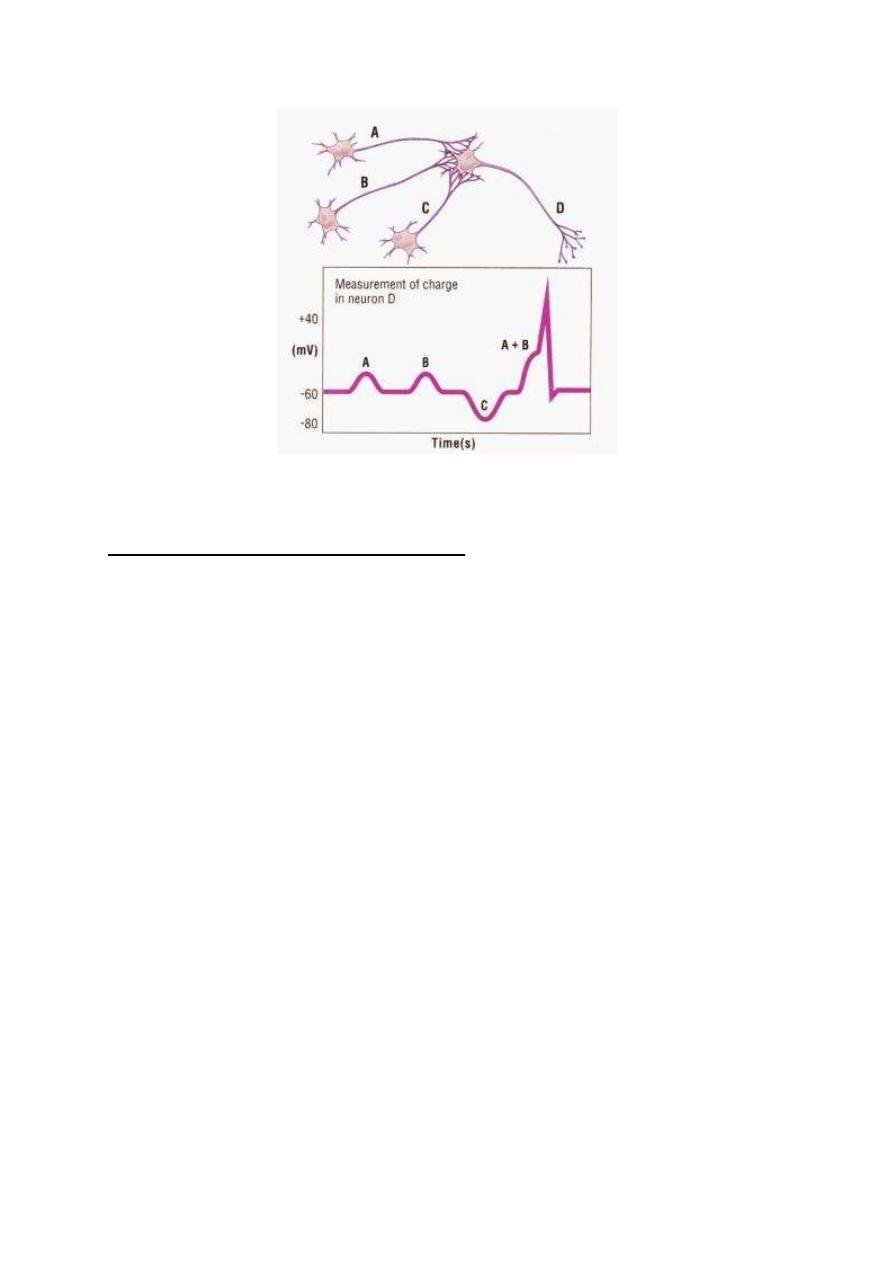
Figure 1-6: The mechanism of spatial summation
THE RESULT OF SUMMATION OF PSPs
An average neuron in the CNS receives about 1000 terminals from
different presynaptic neurons. Some of these terminals are excitatory and
some are inhibitory. Excitatory and inhibitory postsynaptic potentials
may occur at the same time. Summation of PSPs results in one of two
conditions in the postsynaptic neuron:
1. INHIBITORY STATE
This occurs when the inhibitory input is greater than the excitatory input
so IPSPs are produced and the cell membrane becomes hyperpolarized
and its excitability decreases.
2. EXCITATORY STATE
This occurs when the excitatory input is greater than the inhibitory input so
more EPSPs are produced and the cell membrane becomes depolarized.
When depolarization reaches a critical threshold level (the firing level) a
propagated action potential (nerve impulse) is produced.

The propagated action potential starts at the initial segment of the
neuronal axon not at the soma of the neuron because:
i. The density of the voltage-gated Na
+
channels at the initial
segment is seven times as much as those at the membrane of the
soma. This allows greater and faster Na
+
influx which creates more
rapid depolarization up to the firing level.
ii. The depolarization required to open the voltage gated Na
+
channels at "the initial segment is only +15 mV, whilst the required
depolarization at the soma is +30 mV.
This explains why the axoaxonic synapses are the most effective in
exciting the postsynaptic neuron. This is because they are the nearest
to the initial segment. The least effective synapses are the
axodendretic synapses.
THE EXCITATORY AND INHIB ITORY NEURONS
The presynaptic neuron is either excitatory or inhibitory to the
postsynaptic neurons. This is because neurons can release only one type
of transmitter which is either excitatory or inhibitory to the postsynaptic
neuron. A cotransmitter may be released with the primary
transmitter. It is, however, always a potentiator of the primary
one.
PRESYNAPTIC AND POSTSYNAPTIC INHIBITION
Transmission of impulses across the synapse can be inhibited or
blocked in two ways (fig. 1-6):

1. PRESYNAPTIC INHIBITION
Presynaptic inhibition is the inhibition of synaptic transmission by
inhibiting the release of the transmitter from the presynaptic nerve
terminal. It can be induced by certain drugs (botulinum, toxin) or by
certain inhibitory neurons (attenuators) which synapse on presynaptic
terminals. The attenuator neuron inhibits the synapsin phosphorylase
enzyme of the excitatory neuron so the transmitter vesicles remain
attached to the cytoskeleton and no release of the transmitter.
Figure 1-7: Presynaptic and postsynaptic inhibition.
2. POSTSYNAPTIC INHIBITION
Postsynaptic inhibition is inhibition of synaptic transmission by
induction of an inhibitory state in the postsynaptic neuron. It is induced
either by drugs or by inhibitory neurons.

Physiology
Dr. Basim Mohamad Alwan Lecture (2)
PROPERTIES OF SYNAPTIC TRANSMISSION
Transmission of signals across the synapses is characterized by:
1. FORWARD DIRECTION
Transmission in synapses is unidirectional, i.e. from the presynaptic to
the postsynaptic neuron, not the reverse. This is because the postsynaptic
neuron cannot release a chemical transmitter at the synapse. So, the
synapse acts as a unidirectional "valve" to keep the flow of signals
between neurons always in the right direction.
2. SYNAPTIC DELAY
When an impulse reaches a nerve terminal, it takes a delay time of
0.5-1.0 ms to pass across the synapse to the postsynaptic neuron. This
time is taken for the release of the chemical transmitter, its diffusion in
the synaptic extracellular fluid, activation of receptors, induction and
summation of postsynaptic potentials.
3. SYNAPTIC AFTERDISCHARGE
After discharge is the persistence of output signals after stoppage of
the input signals. Synaptic afterdischarge occurs at some synapses
because of the delay of inactivation of the chemical transmitter. So, an
impulse conducted by a presynaptic neuron may produce more than,
one impulse in the postsynaptic neuron. The duration of the synaptic
afterdischarge is longer if the chemical transmitter released by the

presynaptic neuron is a long acting one (substance P).
4. FATIGUE
Fatigue is the decline in response caused by prolonged activity. For a
synapse, fatigue is the decline in the response of the postsynaptic neuron
after a long period of high frequency stimulation of the synapse (> 60
Hz). It is manifested by prolongation of the synaptic delay, then failure to
transmit some or all of the impulses across the synapse.
The synapse is an early site of fatigue in the reflex arc and the fatigue of
the neural synapses is caused by:
i. Exhaustion of the chemical transmitter in the presynaptic terminals
which is the main cause.
ii. Inactivation of some postsynaptic receptors due to accumulation of
metabolites.
iii. Marked increase of the intracellular Ca
2+
in the postsynaptic
neuron. This high Ca
2+
level opens K
+
channels so K
+
efflux and
hyperpolarization of the postsynaptic membrane decreasing the
excitability of postsynaptic neuron.
Fatigue is a protective mechanism against excess neuronal activity;
e.g. fatigue is the most important means by which the excess
excitability of an epileptic circuit is cut off and stopped. This leads to
spontaneous ending of the epileptic fit (normal protective mechanism).
5. SYNAPTIC POTENTIATION (FACILITATION)
This is an increase in the postsynaptic response caused by previous
presynaptic stimulation.
It may be a short-term or a long-term potentiation.

A. SHORT TERM (POST TETANIC) POTENTIATION
This occurs after a short period of low frequency stimulation of the
synapse (< 60 Hz). It is caused by an increase in the intracellular Ca
2+
level in the presynaptic neuron, which increases the release of the
transmitter. Short-term potentiation lasts for few seconds up to few
minutes.
B. LONG TERM POTENTIATION (LTP)
This occurs after a short period of high frequency stimulation (>60 Hz).
LTP is caused by the release of arachidonic acid from the postsynaptic
neuron which acts on the presynaptic neuron to release more of the
transmitter (Glutamate).
Long-term potentiation occurs in several parts of the CNS, particularly
in the hippocampus and it plays an important role in memory and
learning.
6. SYNAPTIC DEPRESSION (HABITUATION)
Habituation is the gradual decrease in the postsynaptic response when
stimulation of the presynaptic neuron is frequently repeated. With
complete habituation, the postsynaptic response may disappear
altogether.
Habituation is due to inactivation of Ca
2+
channels in the presynaptic
neuron which decrease in intracellular Ca
2+
so release of smaller
amount of transmitter from the presynaptic terminals. The cause of
this, inactivation is unknown.
Habituation could be short-term or long-term depending on how many
times the stimulus is applied. It is an important mechanism of learning,
as it enables the subject to ignore insignificant stimuli.

Habituation of synapses is different from adaptation which occurs in
excitable tissues. Adaptation is the decline in response to a constant
maintained stimulus.
7. SENSITIZATION
Sensitization of a synapse is the potentiation of the postsynaptic
response to a certain stimulus by coupling the stimulus to another
intense (usually painful) stimulus (fig.2-1).
The terminal which conducts the intense or painful stimulus is called
a facilitator terminal. It relays on the presynaptic sensory terminal.
The facilitator terminal stimulates the presynaptic sensory terminal
lead to prolonged action potential in the sensory terminal and more
Ca
2+
influx into the sensory terminal so release of more transmitter and
potentiated postsynaptic response result.
Sensitization is an important mechanism in memory and learning.
Figure 2-2: The mechanism of synaptic sensitization.
8. EFFECT OF pH
Alkalosis enhances synaptic transmission. A rise of arterial blood pH
from 7.4 to 7.8 leads to increased cerebral excitability and convulsions.
Acidosis depresses synaptic transmission. Breathing of air with high

C0
2
level will lead to hypercapnea and acidosis and then depression of
synaptic transmission in the brain resulting in drowsiness and sleep or
even anesthesia. A drop of arterial pH down to 7.0 produces coma
because of failure of synaptic transmission between various neurons in
the brain.
9. EFFECT OF HYPOXIA
Hypoxia depresses synaptic transmission and prolongs reflex time due
to accumulation of acidic metabolites.
10. EFFECT OF DRUGS
Caffeine, theophylline and theobromine which are found in coffee,
tea enhance synaptic transmission. They increase neuronal excitability
by lowering the threshold of excitation.
Strychnine enhances synaptic transmission by blocking the action of
central inhibitory transmitters (e.g. glycine).
Hypnotics and anesthetics depress synaptic transmission by
decreasing neuronal excitability. They stabilize the cell membrane by
increasing the resting membrane potential (hyperpolarization).
PROCESSING OF SIGNALS IN THE CNS
Nerve signals (impulses) enter the CNS to be directed to various
neuronal pools
(collection of neurons). In the neuronal pools, input
signals are processed, and output signals emerge out to proceed to
specific destinations.
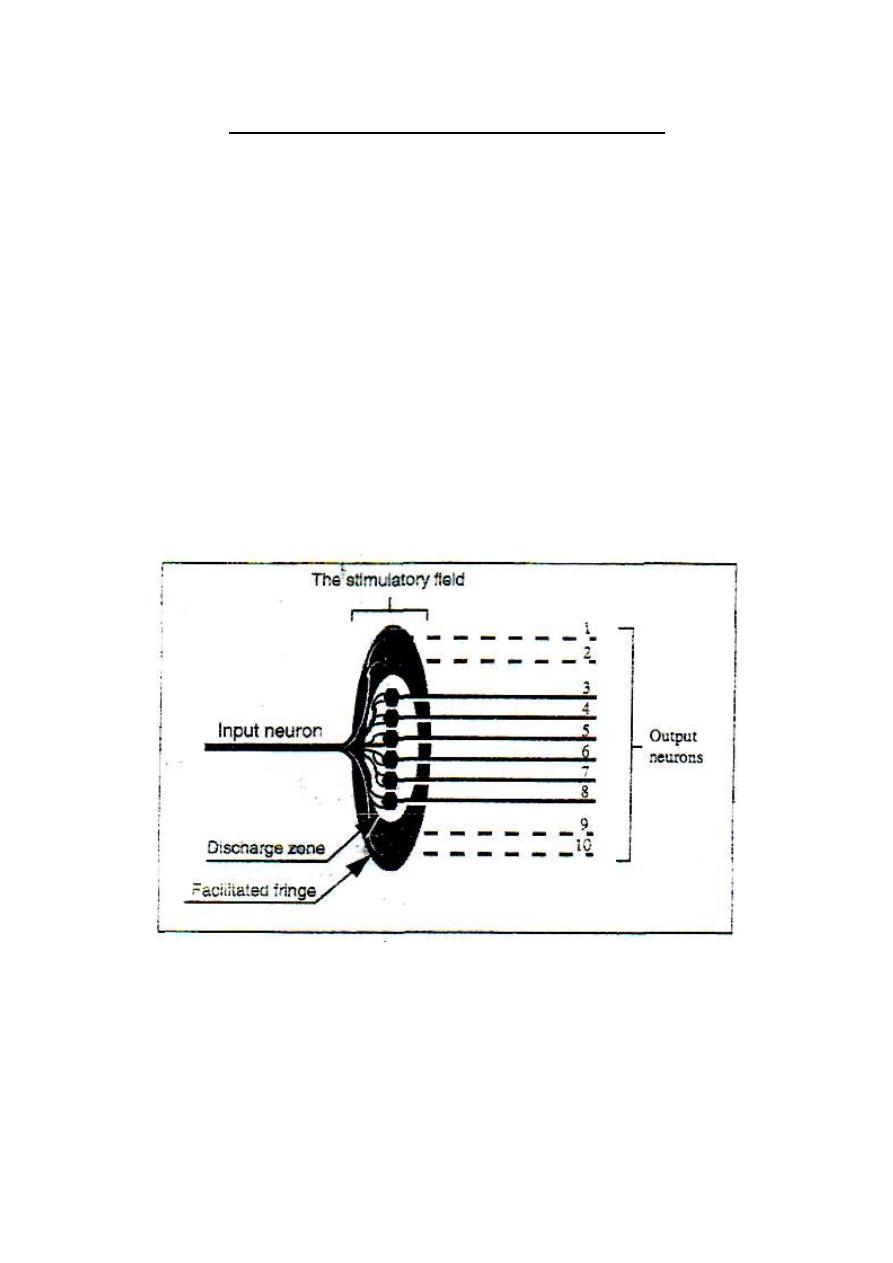
THE DISCHARGE ZONE AND THE FACILITATED FRINGE
(THE LIMINAL ZONE AND THE SUBLIMINAL FRINGE)
When an impulse in an excitatory input neuron reaches the neuronal pool, it
stimulates a group of neurons which form the "stimulatory field'' of this
neuron (fig.2-3).
Figure 2-3: The stimulatory field of an input neuron.
At the middle of the field, stimulation reaches a liminal level
(threshold) and the neurons in this zone discharge impulse. The zone
where neurons discharge impulse is called the discharge zone or the

liminal zone of the input neuron.
Around the discharge zone, there is a circular zone (a fringe) in which
the neurons are only facilitated without reaching the
liminal
firing
level. This zone is called "the facilitated fringe" or "the subliminal
fringe" of the input neuron.
An impulse in an inhibitory input neuron produces an "inhibitory
field'' with maximum inhibition at its center.
FORMS OF SIGNAL PROCESSING IN THE NEURONAL POOLS
Signal processing in the neuronal pools takes one of the following
forms:
[I] Convergence.
[II] Divergence.
[III] Prolongation.
[IV] Shortening.
[V] Sharpening.
[I] CONVERGENCE OF SIGNALS
Convergence is the direction of signals from several input neurons to
excite a single output neuron. There are two main types of convergence
in the neuronal pools.
1. CONVERGENCE FROM A SINGLE SOURCE
This is important because no neuron can be excited by a single input
terminal. So, convergence must occur on neurons to excite those
neurons (fig.2-4). The spatial summation of postsynaptic potentials
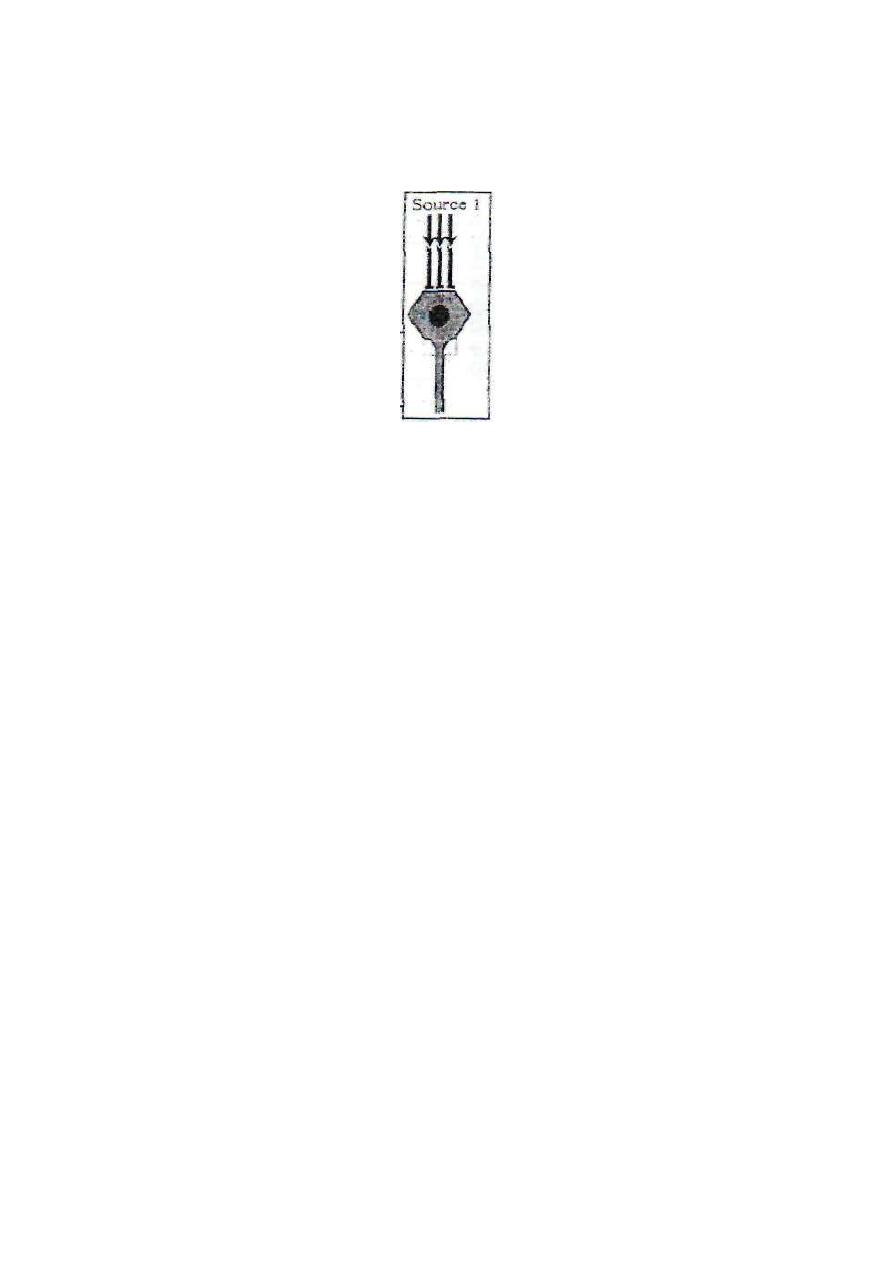
from the multiple input terminals builds up a threshold membrane
potential to excite the neuron.
Figure 2-4: convergence from single
source
2. CONVERGENCE FROM .MULTIPLE SOURCES
This is important because it enables neurons of the neuronal pool to
receive signals from different sources (fig. 2-5).
The effect produced will be the resultant of all the inputs whether
excitatory or inhibitory; e.g. motor neurons of the ventral horn of the
spinal gray matter receive inputs from the pyramidal and extra
pyramidal tracts and from the afferent fibers of the stretch reflex and
several intermediate neurons. All these input neurons influence the
contraction and relaxation of the skeletal muscles.
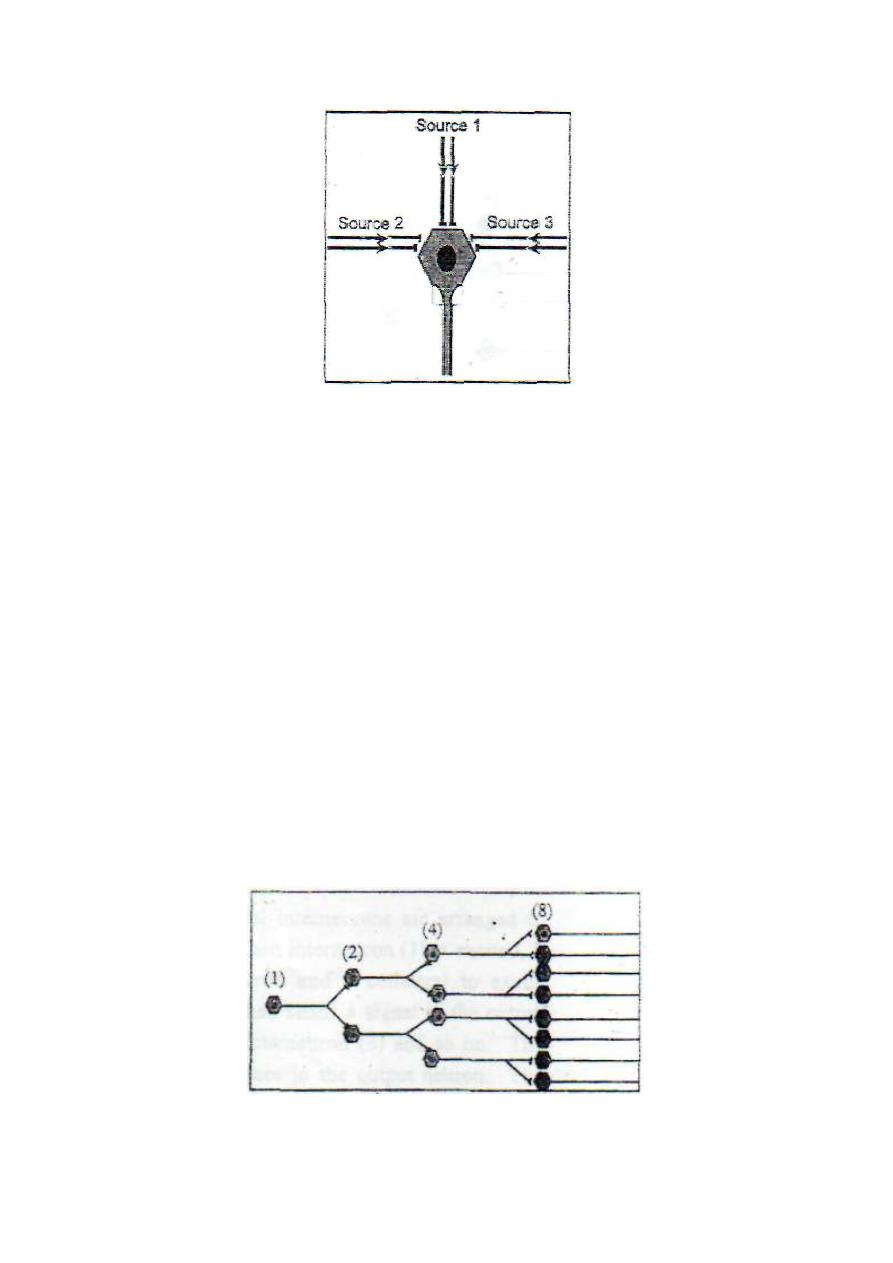
Figure 2-5: Convergence from multiple sources.
[II] DIVERGENCE OF SIGNALS
Divergence is the spread of a signal from one input neuron into more
than one output neuron. There are two main types of divergence in the
neuronal pools:
1. DIVERGENCE IN THE SAME BATHWAY
This leads to spread of the signal into an increasing number of neurons
as it passes from one order of neurons into another (fig. 2-6). It may be
called an "amplifying divergence". It occurs, for example, in the
pyramidal tract where a single pyramidal neuron in the motor cerebral
cortex can excite up to 10,000 muscle fibers.
Figure 2 - 6: Amplifying divergence.

2. DIVERGENCE INTO MULTIPLE PATHWAYS
This leads to spread of the signal into two or more separate directions
from the pool (fig .2-7). It may be called a "diversifying divergence".
It occurs, for example, in the paleospinothalamic tract where
some signals proceed directly to the thalamus and others enter the
spinoreticular tract.
Figure 2-7: Diversifying divergence.
[III] PROLONGATION OF SIGNALS (AFTERDISCHARGE)
Afterdischarge is the persistence of output signals after stoppage of the
input signals. This is possible through the following mechanisms:
1. SYNAPTIC AFTERDISCHARGE:
2. OPEN-OHAIN CIRCUITS
These are circuits in which several interneuron's are arranged to form
an open circuit (fig. 2-7). When interneuron (1) is excited, it sends a
signal to the output neuron and collateral to excite interneuron (2).

Interneuron (2) then sends a signal to the output neuron and collateral
to excite interneuron (3) and so on. The result will be a barrage
(train of
impulses follows each other)
of impulses in the output neuron. The
interneurons of the open-chain circuits are called "the interneuronal
barrages.
Figure 2 - 8: An open chain circuit with interneuronal barrages
3. CLOSED {REVERBERATING} CIRCUITS
These are circuits in which the output neuron is repeatedly activated
through a closed circuit of interneurons (fig.2-8). When interneuron (1)
is excited, it sends an impulse to the output neuron and collateral to
excite interneuron (2). Interneuron (2) then re-excites interneuron (1),
and so on.
Figure 2 - 8: A closed chain (reverberating) circuit of neurons.

Reverberating circuits can be facilitated or inhibited by other input fibers.
When facilitated the frequency and duration of discharge in output fibers
increase. When inhibited, the frequency and duration decrease.
The cycle of reverberation stops by either fatigue of synapses or
inhibition by other input fibers. The frequency and duration of
discharge from a reverberating circuit depends on the number of
neurons (i.e. the number of synapses) in the circuit. The larger the
number the lower is the frequency and the longer is the duration.
RHYTHMIC SIGNAL OUTPUT
Certain centers in the CNS produce rhythmic signals, e.g. the
respiratory center in the brainstem. This rhythmic signal output is
caused by reverberating circuits involving a large number of
interneurons. The large number of synapses slows down the frequency
of output discharge and delays fatigue of synapses.
[IV] SHORTENING OF SIGNALS
Shortening of signals means suppression of afterdischarge in the output
neurons. This is done by either feedback or feed forward inhibition.
1. FEEDBACK INHIBITION
This occurs when an excitatory neuron stimulates an inhibitory neuron
then the inhibitory neuron turns back to inhibit the initial excitatory
neuron. In this case, stimulation of a neuron results in feedback
inhibition of the same neuron to shorten the duration of discharge and
prevent any afterdischarge. This occurs, for example, with the spinal
motor neurons (the ventral horn cells). Each spinal motor neuron

regularly gives off a collateral branch which synapses with an inhibitory
interneuron called "Renshaw cell".
Renshaw cell sends inhibitory signals to the cell body of the original
spinal motor neuron (feedback inhibition). The inhibition of the original
motor neuron suppresses any synaptic afterdischarge to prevent
undesired prolonged activity of the motor nerve.
2. FEEDFORWARD INHIBITION
This occurs when an input neuron stimulates an output neuron plus an
inhibitory interneuron then the inhibitory interneuron inhibits the output
neuron (fig. 2-9). In this case, stimulation of an input neuron results in
stimulation then rapid inhibition of the output neuron. This prevents any
undesired prolonged discharge from the output neuron.
Feed forward inhibition occurs in the cerebellum where a single input
neuron stimulates an output neuron and a Purkinje cell.
The Purkinje cell then inhibits the output neuron, cutting short any
undesired afterdischarge.
Figure 2 - 9: Feedforward inhibition.
[V] SHARPENING OF SIGNALS
Sharpening of signals means the limitation of signals to the target
neurons only. This requires the inhibition of any undesired activity in
the nearby neurons. This is achieved by either lateral or reciprocal
inhibition mechanisms.

1. LATERAL INHIBITION
Lateral inhibition occurs when a neuron sends collaterals to inhibit the
nearby neurons through intermediate inhibitory neurons. This helps to
focus the activity to the original neuron and eliminate any undesired
discharge from the nearby neurons. The function of Renshaw" cell
shows an example of both feedback inhibition (of the original motor
neuron) and lateral inhibition (of the nearby neurons).
Lateral inhibition occurs also in sensory neurons. Sensory fibers
conducting touch (scratching) laterally inhibit itch and pain conducting
fibers at the dorsal horn of the spinal gray matter. In this way scratching
relieves itch and pain sensations.
2. RECIPROCAL INHIBITION
In reciprocal inhibition the activation of one output neuron is
accompanied by simultaneous inhibition of another output neuron. This
form of inhibition occurs, for example, during the flexor withdrawal
reflex. In this reflex, contraction of the flexor muscles is accompanied
by concomitant reflex relaxation of the extensor muscles. Reciprocal
inhibition helps to optimize the reflex response by inhibiting any
antagonistic contraction.

Physiology
Dr. Basim Mohamad Alwan Lecture 3
SENSORY NERVES
Sensory nerves (afferent nerves) are the nerves which convey the
sensory information from different parts of the body to the central
nervous system. They make the first order of neurons in the nervous
pathways of all sensations.
All the spinal sensory nerves enter the spinal cord through the dorsal
roots (sensory roots) of the spinal nerves. The cell bodies of these
nerves are in the dorsal root ganglia of the spinal nerves. As these
nerves carry impulses towards the cell bodies they are considered as
long dendrites of the ganglion cells.
BELL-MAGENDI LAW: It states that the dorsal roots of the spinal
nerves are sensory and the ventral roots are motor".
RECEPTIVE FIELD: the spatial region where application of a stimulus causes a
sensory neuron to respond.
receptive fields can overlap
definition applies to higher order neurons, as well as to primary
afferents
SENSORY UNIT: a primary afferent and the receptors that define its receptive
field.
DERMATOME
A dermatome is the area of skin which is supplied by a spinal dorsal
root i.e. it is the cutaneous receptive field of a spinal dorsal root (fig.
4-1). The dermatomes of different roots overlap with each other.
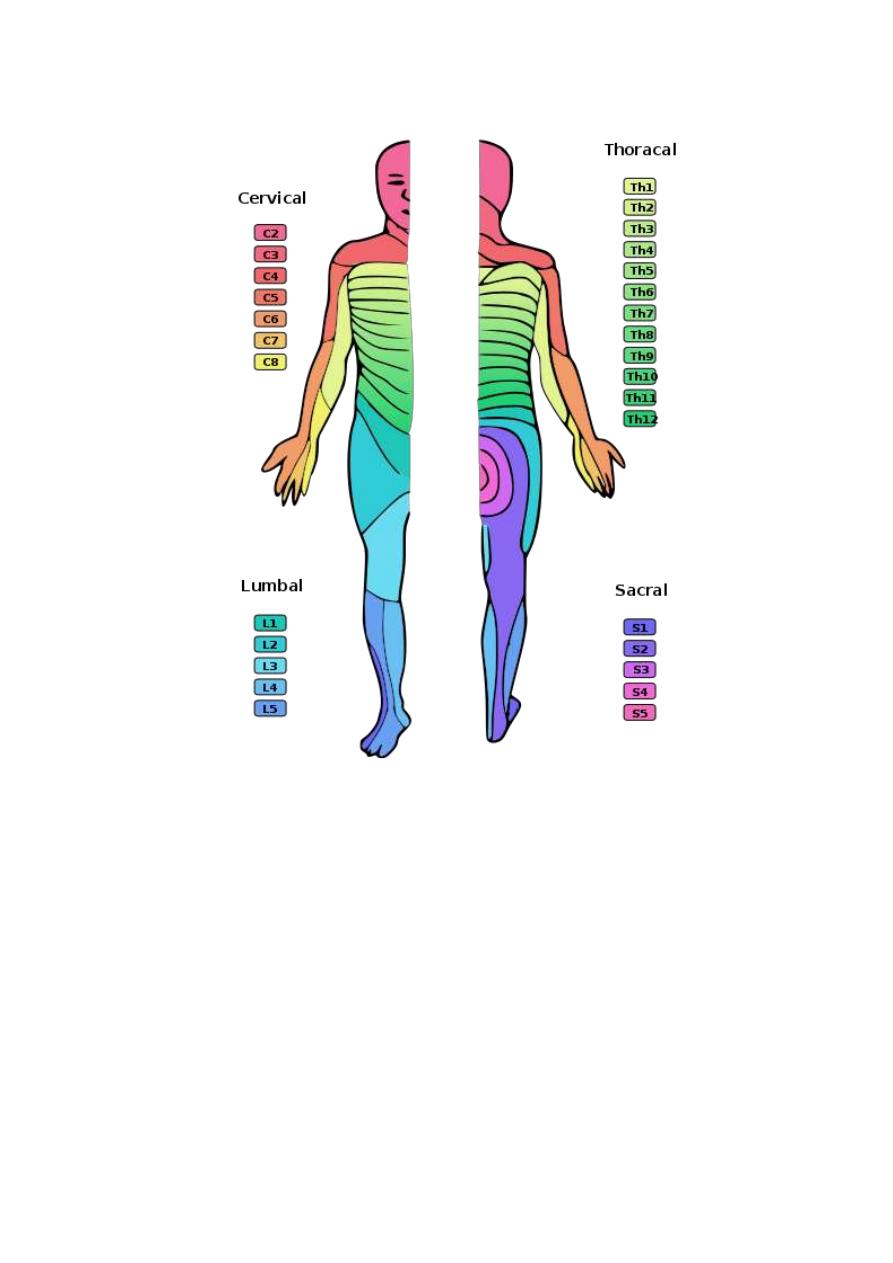
Figure 3 - 1: Dermatomes of the spinal ant. and post. roots
THE DORSAL HORN OF THE SPINAL GRAY MATTER
The
dorsal horn of the spinal gray matter is the site of relay of most of
the afferent sensory fibers. A cross section in the spinal cord shows
several anatomical laminae of the gray matter. Each type of sensory
fibers relays in certain specific laminae as shown in fig. 4-2.
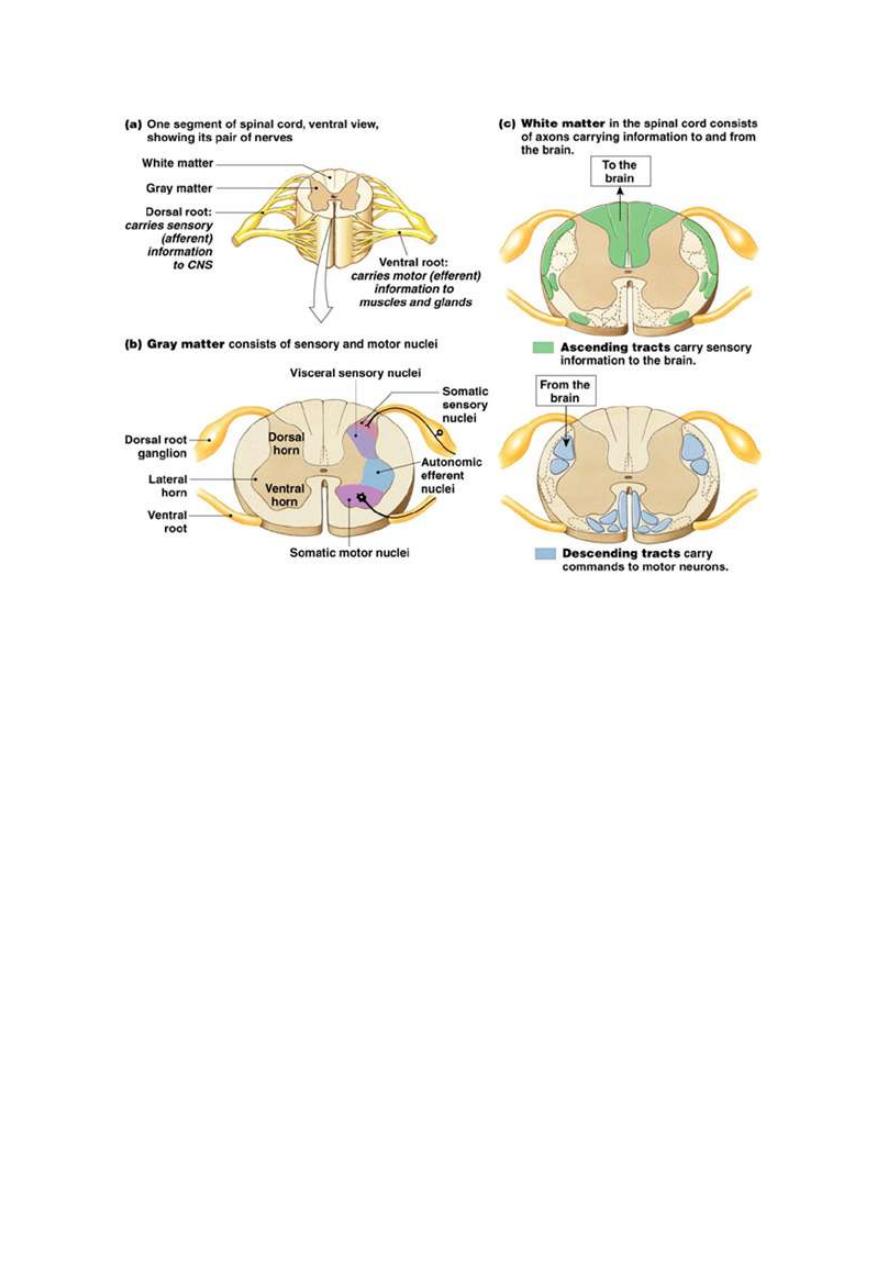
Fig.3-2 the dorsal and ventral horn of SC
THE COURSE OF THE AFFERENT SPINAL NERVE AFTER ENTERING
THE SPINAL CORD
As the afferent spinal nerve fiber enters the spinal cord, it takes one of
the following courses:
1. Ascends or descends for few segments at the tip of the dorsal horn
forming the Lissaur tract before entering into the dorsal horn.
2. Enter the dorsal horn to relay on neurons in different laminae of the
dorsal horn. These neurons are either interneurons (intermediate
neurons) or second order neurons of their sensory pathway.
3. Proceeds in the spinal gray matter to relay on motor neurons in the
ventral horn (the reflex arc of the stretch reflex).
4. Ascends without relay in the dorsal column of the spinal white

matter forming the dorsal column tracts (gracile and cuneate tracts)
terminate in the dorsal column nuclei (graclle and cuneate nuclei) in
the medulla oblongata.
SENSORY RECEP TORS
A sensory receptor is a specialized nerve ending which is sensitive to a
specific type of stimulus and produces a specific type of sensation.
FUNCTIONS OF RECEPTORS
1. Detectors of the type of the stimulus. Each receptor is specialized to
detect a certain type of stimulus, e.g. touch, heat, pain etc.
2. Sensitizers, which lower the threshold of stimulation of nerve
endings.
3. Transducers, which convert the energy of the stimulus into an
electric response, i.e. a membrane potential which generates an action
potential in the afferent nerve.
4. Gauges, which measure the intensity of the stimulus.
Accordingly, it can be concluded that without receptors, the CNS
becomes almost useless.
PROPERTIES OF THE SENSORY RECEPTORS
(A) SPECIFICITY
Each receptor is highly sensitive to a certain type of stimulus which is
called the "adequate stimulus" for this receptor. When the adequate
stimulus is used, the receptor is stimulated by the least amount of
energy. If the receptor is stimulated by a stimulus other than its
adequate stimulus, it needs high energy and still gives its specific type of
sensation; e.g. light receptors in the eye could be stimulated by a strong
mechanical blow on the eye, these results in seeing flashes and stars.
(B) EXCITABILITY
(THE GENERATOR OR THE RECEPTOR POTENTIAL)
When a receptor is stimulated, it increases the permeability of the
membrane of the nerve ending to Na
+
so Na
+
influx and membrane
depolarization is produced. The depolarization of the membrane of the
sensory nerve ending on stimulation of the receptor is called the
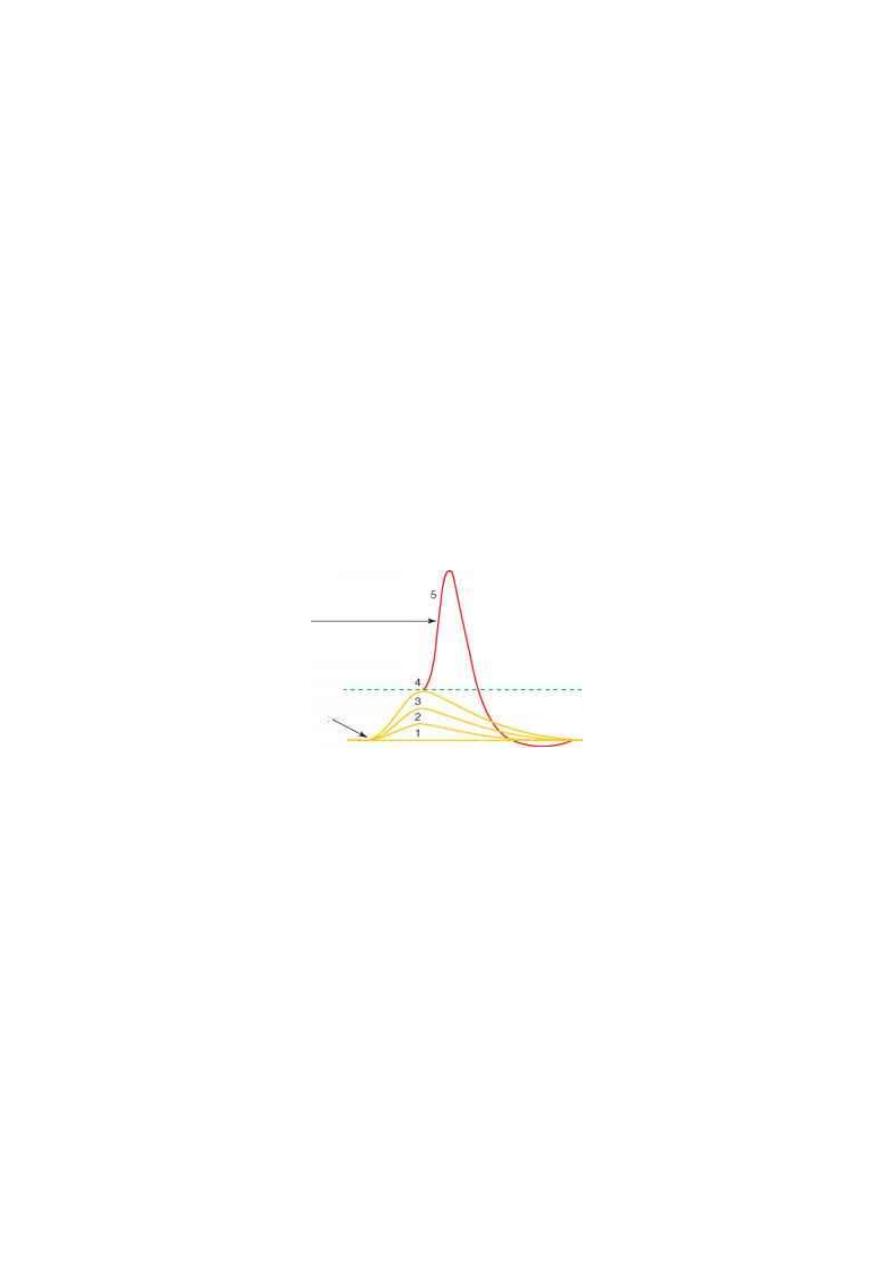
"generator potential" or the "receptor potential" (fig. 3-3). The
generator potential is characterized by:
Figure 3 - 3: The generator potentials produced by 4 increasing intensities of stimuli
(1,2,3,4). When the generator potential reaches the firing level, a full action potential
is produced in the afferent nerve.

The generator potential is characterized by:
1. It does not obey the all or none rule. Its magnitude increases
proportionately with the intensity of the stimulus.
2. It is not followed by a refractory period.
3. It has a long duration (more than 5 ms). So, it can be temporally
summated.
4. When it reaches a threshold value, it activates the first node of
Ranvier of the afferent nerve to generate an action potential in the
afferent nerve.
Figure 3 - 4: The relationship between the magnitude of the generator potential and
the frequency of discharge of impulses in its afferent nerve.
5. It is not blocked by local anesthetics. Local anesthetics prevent the
development of the action potential at the first node of Ranvier but do
not prevent the development of the generator potential.
When the generator potential exceeds the threshold value, the
frequency of discharge of impulses in the sensory nerve becomes
directly proportionate with the amplitude of the generator potential
(fig. 3-3).
(C) DISCHARGE OF IMPULSE
The response of a receptor to a change in the intensity of the stimulus is
by changing the "frequency" of impulses generated in the afferent
nerve. That is why the response of receptors: to a change in stimulus
intensity is called a "frequency modulated response" or "FM
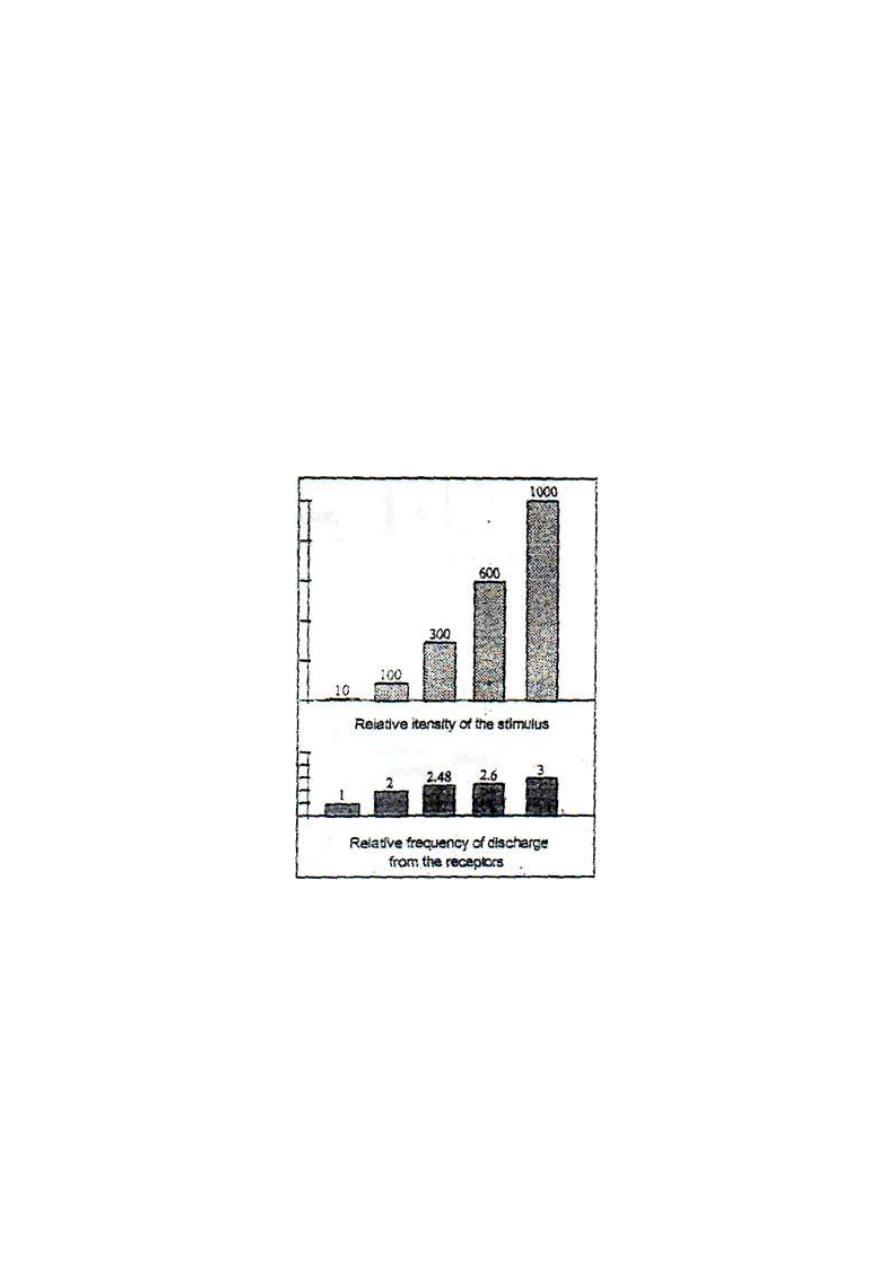
response". Weber-Fechner law states that "The frequency of discharge
of impulses from a receptor is directly proportionate with the log
intensity of stimulus.
Figure 3 - 5: The compression function of receptors
.
For example, a hundred-fold increase in the intensity of the stimulus
leads to only twofold increase in the frequency of discharge of impulses
from the receptor. The modulation of large changes in the intensity of the
stimulus to small changes in the frequency of impulses is referred to as
"the compression function" of the receptor (fig.3-5). This function
enables the receptor to respond to, and discriminate, a wide range of
stimulus intensity, e.g. sound receptors can inform the CNS of sound
intensity range of one to ten billion-folds.
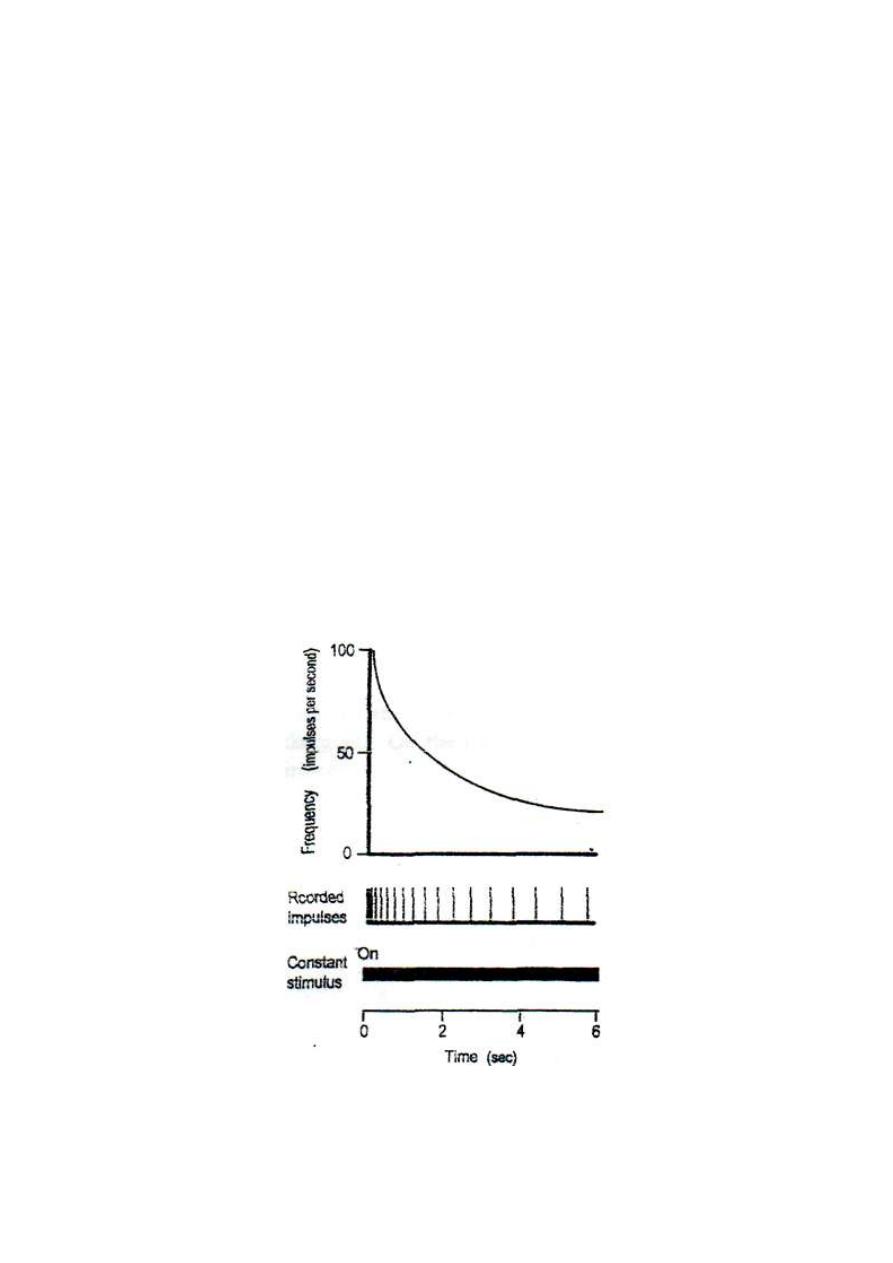
(D) ADAPTATION OF RECEPTORS
Adaptation is the decline in response to a constant maintained stimulus.
If a constant maintained stimulus is applied to a receptor, the frequency
of discharge of impulses in its afferent nerve declines with time. The
rate of decline depends on the type of receptor (fig. 3-6).
The sensory receptors are classified according to their rate of
adaptation, into three types:
Figure 3-6: Adaptation of receptors

1. Rapidly adapting receptors (phasic receptors), e.g. touch and hair touch
receptors.
2. Moderately adapting receptors e.g. thermoreceptors in the
temperature rangebetween 20-40°C.
3. Slowly adapting receptors (tonic receptors), e.g. muscle spindles
and Golgi tendon organs.
The rate of adaptation of each type of receptors fits its function. E.g.
touch receptors adapt rapidly, so after putting clothes on; it would be
irritating to feel the touch of clothes all the time. So, touch receptors
adapt rapidly and stop discharging. On the other hand, muscle spindles
send continuous proprioceptive signals to the CNS which maintain
body posture and equilibrium which are needed all the time, so they
adapt very slowly.
Figure 3 - 7: rate of adaptation of different types of receptors.
MECHANISM OF ADAPTATION OF RECEPTORS
The following factors contribute to the process of adaptation of
receptors:
1. Gradual inactivation (closure) of some Na
+
channels in the
membrane of the nerve ending so membrane depolarization becomes
difficult.
2. Dissipation of some of the stimulus energy to the surrounding tissues
i.e. redistribution of the energy of the stimulus.
3. Decreased sensitivity of the node of Ranvier.
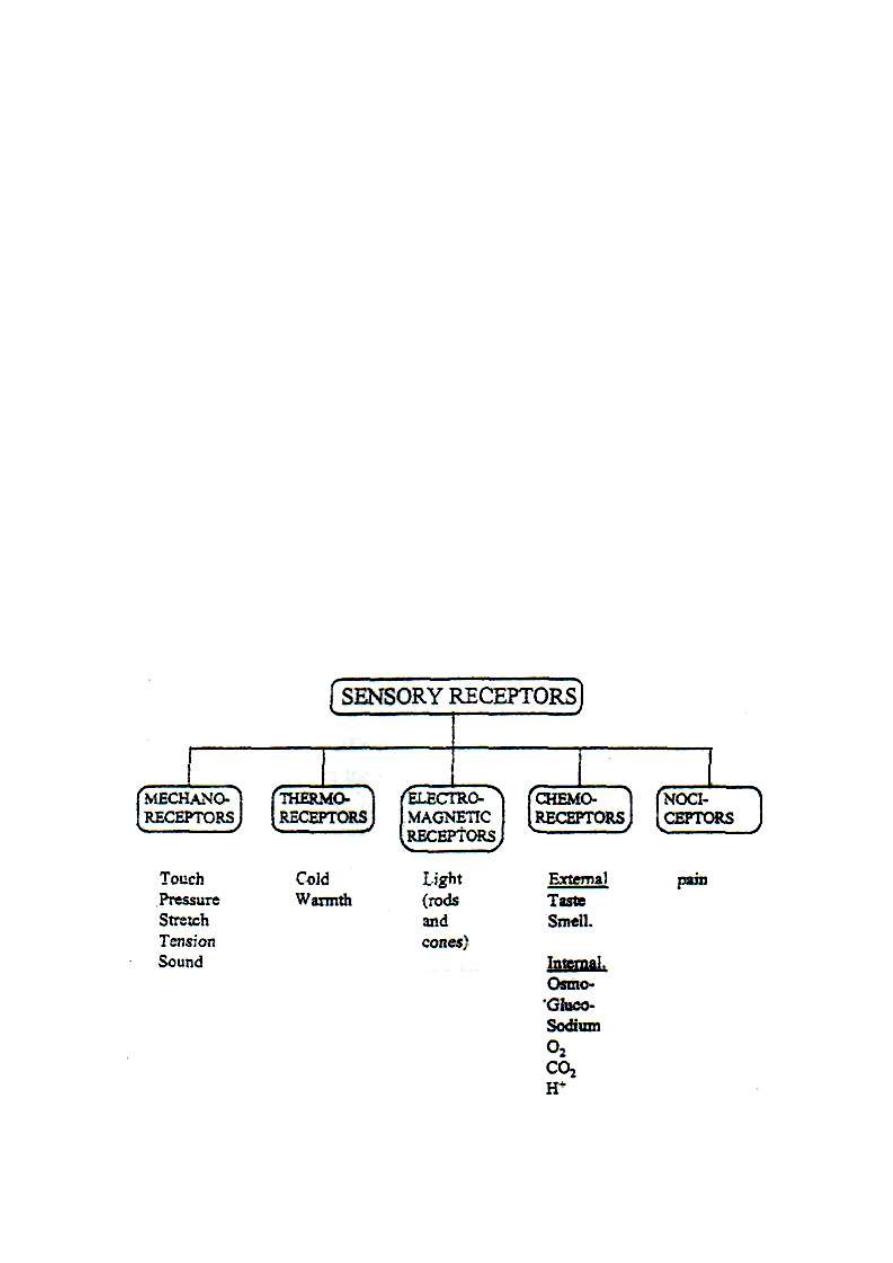
CLASSIFICATION OF RECEPTORS ACCORDING TO THEIR ADEQUATE
STIMULUS
According to the type of their adequate stimulus, receptors are
classified into 5 main categories as shown in fig. 3-8.
Figure 3-8: The five types of receptors according to their adequate stimuli.

CODING OF SENSORY INFORMATION
All stimuli are transduced by the sensory receptors into nerve impulses
in the afferent nerves. These impulses reach the brain acting as code
signals specific for each sensation. The brain then deciphers these code
signals and identifies the modality (type), the locality and the
intensity of the stimulus.
1. CODING OF THE MODALITY OF THE STIMULUS
Each sensory pathway from the receptor up to the final sensory neuron in
the brain is specialized to serve a specific sensory modality.
Stimulation of any point along the course of any sensory pathway
produces its specific sensation. The reservation of a specific sensory
pathway to serve a specific type of stimulus is referred to as "the
labeled line principle"; i.e., the whole line from the receptor to the
final sensory neuron in the brain is "labeled" for conducting impulses of
a specific sensation.
So, the coding of a stimulus modality is by sending the impulses
through its specific sensory pathway (i.e. its labeled line).
LABELED LINE PRINCIPLE
(MULLER'S LAW OF SPECIFIC NERVOUS ENERGY)
This law states that "stimulation of any point along the course of a
sensory pathway produces the specific sensation served by this
pathway regardless of the type of the stimulus used",

2. CODING OF THE LOCALITY OF THE STIMULUS
Each locality sends sensory impulses to the brain via a specific sensory
pathway. Stimulation of any point along the course of this pathway
produces a sensation felt at this specific locality. So, the locality of the
stimulus is also coded by the specific sensory pathway which serves
this locality. This is called the "law of projection". It explains the
phantom limb phenomenon which occurs in amputees.
THE PHANTOM LIMB PHENOMENON
This is the false sensation from a limb when the limb does not really exist. It
occurs in amputees who complain of pain, touch, itching or pressure
sensation felt in the absent limb. This false sensation is due to irritation of
the cut ends of the afferent nerves of the amputated limb which send
impulses up to the brain. The brain projects the sensation on to the absent
limb as if it were existing.
3. CODING OF THE INTENSITY OF THE STIMULUS
The intensity of the stimulus is coded by two factors:
a.
The frequency of discharge from the sensory receptor. The
frequency is proportionate with the log intensity of the stimulus
(Weber Feschner law)
b.
The number of activated receptors. A stronger stimulus activates
more receptors. The activation of more receptors by stronger stimuli is
called "recruitment of receptors".

Physiology
Dr. Basim Mohamad Alwan Lecture (4)
SOMATIC SENSATIONS
Somatic sensations are sensations which are conducted by the somatic nerves and
come to conscious perception. They are classified into three categories;
[I] MECHANORECEPT1VE SENSATIONS
These are sensations produced by mechanical stimuli. They include senses of
touch (crude and fine), tickle and itch, texture of material, vibration, pressure,
stereognosis, muscle tension and proprioception.
[II] THERMORECEPTIVE SENSATIONS
These are sensations produced by thermal stimuli. They include warmth and cold
sensations.
[III] PAIN SENSATION
MECHANORECEPT1VE SENSATIONS
These sensations produced by mechanical stimuli which stimulate
mechanoreceptors. It is a group of sensations which comprises several sensory
modalities.
MECHANORECEPTORS
Mechanoreceptors are receptors which are especially sensitive to mechanical
stimuli. They are classified into cutaneous and deep mechanoreceptors as
follows:
1. CUTANEOUS MECHANORECEPTORS
Cutaneous mechanoreceptors (fig- 4-1) are of three types:
i. Naked nerve endings; e.g. the free nerve endings and the basket endings

around hair follicles.
ii. Expanded nerve endings; e.g. Merkel discs and Ruffini corpuscles.
iii. Encapsulated nerve endings; e.g. Pacinian corpuscles and Meissner
corpuscles.
Figure 4 - 1: Cutaneous mechanoreceptors. A: free nerve endings, B: Basket endings
around hair follicle, C: Merkel discs, D: Ruffini ending, E: Pacinian corpuscle, E: Meissner
corpuscle.
These receptors differ in their excitability, rate of adaptation, and the ability to
respond to repetitive stimuli. They are present in the skin all over the body, but
highly condensed in the finger tips and lips which are highly sensitive to touch.
[II] DEEP MECHANORSCEPTORS
These receptors are of two types:
i. Muscle spindles: which are stretch receptors found in the fleshy part of skeletal
muscles?
ii. Golgi tendon organs: which are tension receptors found in the tendons of the
skeletal muscles.
MECHANORECEPTIVE SENSATIONS
1. TOUCH (TACTILE) SENSATION
Touch is the cutaneous sensation produced by light mechanical stimuli. There are
two types of touch sensation;
Crude touch and fine touch.

(A) CRUDE TOUGH
This is touch sensation without accurate identification of the locality or the
number of stimuli.
TICKLE AND ITCH
Tickle is a sensation produced by mild tactile stimulation of certain areas of skin,
usually leading to reflex involuntary laughter.
Itch is a sensation of skin irritation which leads to the desire for scratching of the
skin (the scratch reflex). It is produced by either a moving tactile stimulus a
moving flea or by substances released in the skin as histamine. The powder of
cowhage plant produces itch by releasing an itch-producing substance in the skin.
The receptors for tickle and itch sensation are specialized, highly sensitive,
rapidly adapting free naked nerve endings. They are found exclusively in the
superficial layers of the skin. Tickle and itch signals are transmitted by the slowly
conducting type IV nerve fibers.
Scratching relieves itch by removing the irritating stimulus and by presynaptic
lateral inhibition of the central terminals of the primary itch-conducting fibers.
Nerve fibers carving scratch signals send collaterals inside the spinal cord to
inhibit fibers of itch sensation by presynaptic inhibition mechanism. So, scratch
conducting fibers inhibit the itch conducting fibers by lateral presynaptic
inhibition.
(B) FINE TOUCH
This is touch sensation with accurate identification of the locality and the number
of stimuli. Fine touch sensation is classified into senses of tactile localization
and tactile discrimination.
(a). TACTILE LOCALIZATION
It is the ability to identify the point where the stimulus is applied. It is tested by
touching the skin of blind-folded subject by the tip of a blunt-pointed object, then
asking the subject to open his eyes and point out the site where he was touched.

The acuity of this sense is inversely proportionate to the distance of error in
localizing the stimulus.
(b). TACTILE DISCRIMINATION
It is the ability to identify two tactile stimuli applied simultaneously as two separate
points of contact. It is tested by the compasses test or using discriminator. The two
blunt ends of test compasses are applied to the skin of a blind-folded subject. The
distance between the two ends of the compasses is increased step by step until the
subject feels the two ends of the compass as two separate points. Acuity of this
sensation is inversely proportionate to the two-point threshold which is the
minimal distance at which the two stimuli are felt as two separate points.
Figure 4-2: Two-point discrimination; a: The compasses test, b: Two-point threshold of different
parts of the skin.
Tactile discrimination is not equally developed in different parts of the skin (fig.
4-2). It is highly developed in the tips of the fingers and tongue (two-point

threshold = 2-3 mm) but poorly developed on the back of the trunk (threshold =
65 mm).
For two-point discrimination to occur, impulses from the two points on the skin
should reach two separate final sensory neurons in the sensory cerebral cortex.
Accordingly, there are three conditions which favor the development of high
degree of tactile discrimination in a skin area. These conditions are:
i. High density of touch receptors.
ii. Minimal convergence in the sensory pathway.
iii. Large area of sensory cortical representation in the sensory cortex; i.e. a
large number of final sensory neurons.
These conditions are all found in the skin of the lips and fingertips.
TEXTURE OF MATERIAL SENSATION
This is the ability to identify the substance from which a textile is made; e.g. silk,
wool, cotton, synthetic material ...etc., by touching and without seeing it. It is a
special type of fine touch sensation which is conducted by the gracile and cuneaie
pathway.
VIBRATION SENSE
This is a flickering (repetitive) tactile sensation. There are two types of vibration
receptors; the high frequency, rapidly adapting Pacinian corpuscles which
respond to vibration frequency up to 800 Hz, and the low frequency, slowly
adapting Meissner corpuscles which respond to vibration frequency up to 80 Hz.
Vibration sense is tested by putting the base of a vibrating tuning fork on the skin.
A sense of buzzing or thrill is felt. The tuning fork is usually put on a
subcutaneous bony prominence just to magnify the vibrations. Bone itself is
insensitive to vibrations.
Vibration sense is conducted by the gracile and cuneate pathway (the dorsal
column-lemniscal system). In cases of uncontrolled diabetes mellitus or

pernicious anemia, degeneration of the dorsal column occurs. An early sign of
this degeneration is decreased sensitivity or disappearance of the vibration sense.
2. PRESSURE SENSATION
This is the sensation produced by a strong, blunt, static mechanical stimulus.
There are two types of pressure receptors: the rapidly adapting Pacinian
corpuscles, and the slowly adapting Ruffini endings.
Pressure sensation may be divided according to the intensity of the mechanical
stimulus into two types; light pressure (pressure touch) sensed by cutaneous
receptors, and deep pressure sensed by receptors in deeper structures as fasciae
and connective tissues.
The acuity of pressure sensation is tested by applying different weights on a
supported hand of a blind-folded subject, then, the subject is asked to identify
which weight is heavier and which is lighter.
Like touch sensation, there are two types of pressure sensation:
A. CRUDE PRESSURE SENSATION
This is pressure sensation with low ability to discriminate different weights. This
sensation is conducted by the ventral spinothalamic pathway.
B. FINE PRESSURE SENSATION
This is pressure sensation with high ability to discriminate different weights. This
sensation is conducted by the gracile and cuneate pathway.
STEREOGNOSIS
It is the ability to identify objects by handling them without seeing them (e.g. a
key, a coin, a nail ...etc.). This ability depends on touch and pressure sensations
as well as the cortical sensory somatic association area of the parietal lobe
(Brodmann areas 5 and 7). This sensation is conducted by the gracile and
cuneate pathway.

3. MUSCLE TENSION SENSATION
This is the sensation produced by traction on muscle tendons. The receptors are
the Golgi tendon organs. It enables the person to discriminate different weights
by lifting them. It is tested by applying different weights on an unsupported hand
of a blind-folded subject, then, the subject is asked to identify the lighter and the
heavier weight.
4. PROPRIOCEPTIVE SENSATIONS (PROPRIOCEPTION)
Proprioception is the sensation of the position of different parts of the body
relative to each other and the position of the body in space.
Proprioception is divided into two types:
(A) STATIC PROPRIOCEPTION (SENSE OF POSITION)
Static proprioception is the sense of the position of different parts of the body
relative to each other. The receptors are the deep mechanoreceptors (muscle
spindles and Golgi tendon organs. It is tested by putting a limb in an unusual
position and asking the blind-folded subject to put the other limb in a similar
position.
(B) DYNAMIC PROPRIOCEPTION
(SENSE OF MOVEMENT, KINESTHETIC SENSATION OR KINESTHESIA)
Dynamic proprioception is the sense of movement of joints. The receptors are the
Pacinian corpuscles and Golgi tendon organs in ligaments and synovial
membranes of joints. This sense is tested by moving a joint and the blind-folded
subject is asked to tell when the movement begins and when it stops or when the
rate of movement changes.

THERMORECEPTIVE SENSATIONS
Thermoreceptive sensations are those of warmth and cold. There are two types
of specialized thermoreceptors, one is sensitive to warmth and the other is
sensitive to cold. Thermoreceptors are found in the base of the epidermal layer of
the skin. There are warmth sensitive spots on the skin where there are warmth
receptors only and cold sensitive spots where there are cold receptors only. The
number of cold spots on the skin is 4-10 times as many as those of the warmth
spots. The highest density of thermoreceptors is found in the skin of the face and
hands.
Thermoreceptors are also found in the abdominal viscera, the spinal cord and
around great veins. These receptors are concerned mainly with informing the
hypothalamic thermostat of any increase in the body core temperature. They do
not give rise to warmth or cold sensations.
Cutaneous thermoreceptors monitor the temperature of the skin, not that of the
body. An alcoholic drink in cold weather gives a sense of warmth mainly because
it causes cutaneous vasodilation leading to warming of the skin. In this case, the
sense of warmth reflects an increase in the temperature of the skin, not that of the
body. The body temperature might even decrease because of the excessive heat
loss from the skin.
THERMORECEPTORS
There are two types of thermoreceptors:
1. WARMTH RECEPTORS
Warmth receptors are specialized free nerve endings. They are stimulated at
temperatures between 25-50
o
C with maximum frequency of discharge at about
40°C (fig. 4-3). At temperatures lower than 40
o
C the frequency of discharge
decreases and the discharged impulses take the separated impulses pattern
(ungrouped pattern). At temperatures higher than 40°C, the frequency decreases
and the discharged impulses take a grouped pattern, "volley pattern" (fig.
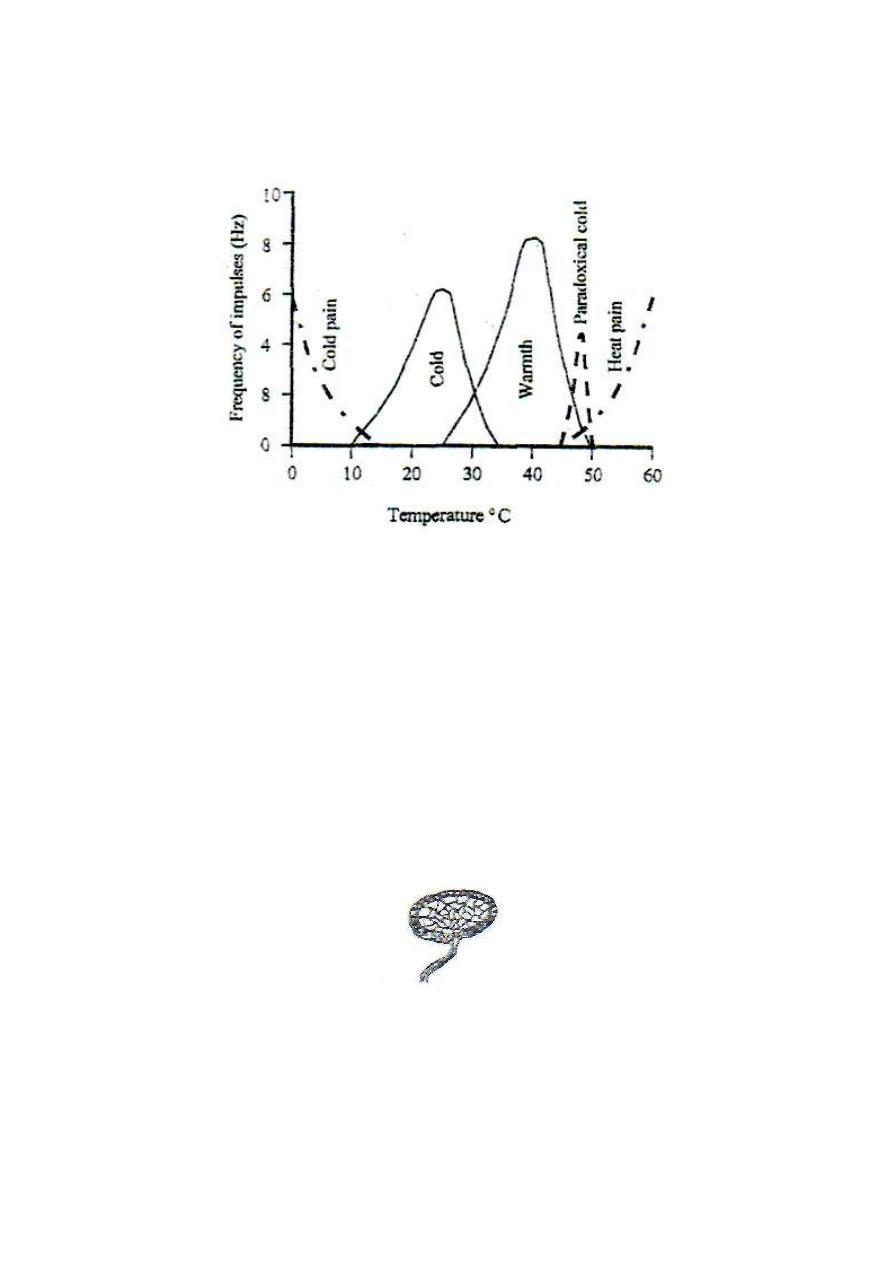
4-5). Warmth sensation is conducted by the thin, type IV fibers.
Figure 7-3: The frequency of discharge from thermoreceptors and thermal pain receptors at
different temperatures.
2. COLD RECEPTORS
Cold receptors are the Krause end-bulbs which are specialized, encapsulated
nerve endings (fig. 4-4). They are stimulated at temperatures between 10-35°C
with maximum frequency of discharge at about 25°C (fig. 7-3). At temperatures
lower than 25°C, the frequency of discharge decreases and the discharged
impulses take the grouped pattern (volley pattern).
Figure 4 - 4: Krause end bulb.
At temperatures higher than 25°C, the frequency of discharge decreases and the
discharged impulses take the ungrouped pattern. Cold sensation is transmitted
by type III nerve fibers.
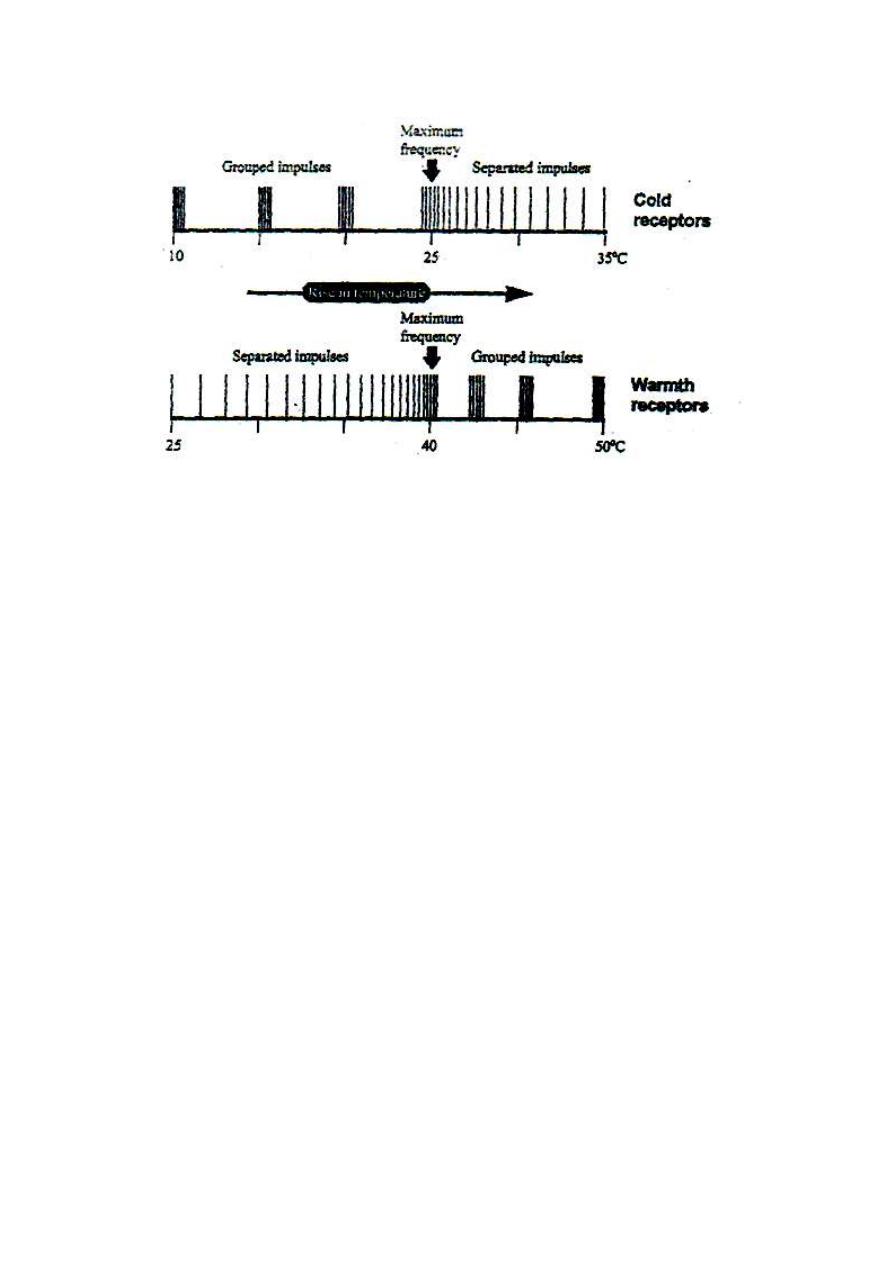
Figure 4-5: The change in
frequency and pattern of discharge from thermoreceptors with the
change of temperature
Thermoreceptors adapt at temperatures between 20-40°C, giving a feeling of
thermeneutrality, but no adaptation occurs outside this range. At temperatures
below 15
o
C or above 45°C, thermosensitive pain receptors are activated giving
rise to pain sensation (cold pain and heat pain). Pain sensation at temperature
ranges of 10-15 and 45-50 is very mild and masked by the senses of extreme cold
or extreme warmth respectively. The sense of thermal pain is clearly perceived at
temperatures below 10 or above 50 °C.
STIMULATION OF THERMORECEPTORS
The effectiveness of a stimulus applied to thermoreceptors depends on two
factors:
1. The absolute temperature: In the temperature range of 10 - 50°C, anything
warmer than the skin is felt warm and anything colder than the skin is felt cold.

2. The rate of change of temperature; i.e. the rate of warming or cooling. A
rapidly changing temperature (rising or falling) is much more effective stimulant
to thermoreceptors than a slowly changing or a steady temperature.
PARADOXICAL COLD SENSATION
Rapid warming of the skin to temperatures between 45-50
o
C gives a transient
false sense of cold (paradoxical cold sensation). This is because at this
temperature range, cold receptors are transiently stimulated (fig. 4-3).
Paradoxical cold, like any cold sensation, produces reflex vasoconstriction and
rise in arterial blood pressure (the cold pressor effect).
Hot water showers are not recommended for cardiac patients because they may
give a sense of cold and then cold pressor effect which lead to rise in arterial blood
pressure and increase in the work load on the heart and heart failure.
NONSPECIFIC THERMORECEPTORS
Some pressure receptors (Ruffini endings) are stimulated also by cold. This
explains why a colder of two otherwise identical weights placed on the hand is felt
heavier than the warmer weight (Weber's illusion). This is because the colder
weight is a stronger stimulant of pressure receptors than the warmer weight
because it makes double stimulation of Ruffini pressure receptors, first by its
weight, and second by its coldness.

Physiology
Dr. Basim Mohamad Alwan Lecture (5)
PAIN
Pain is an unpleasant sensation produced by damage or impending damage of
tissues. It is a specific sensation produced by stimulation of specific pain
receptors (nociceptors). It is not due to overstimulation of other receptors as
mechano or thermoreceptors as was believed before. This is proved by:
1. Pain can be dissociated from other sensations; e.g. in syringomyelia (saccular
dilation of the spinal central canal) pain and temperature sensations disappear
from the affected areas but other sensations persist.
2. Some areas do not contain pain receptors and are insensitive to pain (e.g. the
center of the cornea, the inside of the cheek opposite the second molar tooth),
these areas are sensitive to other sensations.
3. Strong mechanical stimulation of light receptors of the eye (e.g. by a blow on
the eye) produces light sensation from the light receptors (flashes and stars) not
pain.
CHARACTERISTICS OF PAIN SENSATION
Pain has the following characteristic properties
1. Widespread distribution: Pain receptors are present almost everywhere in the
body. It is most abundant in the skin.
2. High threshold of stimulation: The stimuli used to produce pain sensation
(noxious stimuli) are of high energy that causes damage of the tissues.
3. It is produced by a variety of stimuli; mechanical, thermal or chemical.
4. It is a nonadapting sensation; pain sensation persists so long as the noxious
stimulus is applied, the sensation of pain might even increase.

5. It produces prepotent withdrawal reflexes; a prepotent reflex is a reflex that
takes priority over any other reflex occurring at the same time. E.g. swinging of
the arms is an automatic reflex that occurs during walking. If during walking a
noxious stimulus is applied to the arm, swinging stops and the arm is rapidly
withdrawn away from the stimulus.
6. It is the only sensation which produces an unpleasant "affect" without
any previous learning or experience. The pleasantness or unpleasantness of
other sensations are conditioned and depend on previous learning and experience.
Only pain has a "built-in" unpleasant affect. Prefrontal lobotomy (an operation to
cut the deep connections of the frontal lobes with the rest of the brain) abolishes
this affect. After this operation, the patient feels the pain, but it does not bother
him.
7. It can be perceived at the thalamus. The cerebral cortex is not essential for
the perception of the slow type of pain.
PAIN RECEPTORS
There are three types of pain receptors:
1. Mechanosensitive pain receptors: Which are stimulated by strong
mechanical stimuli?
2. Thermosensitive pain receptors: Which are stimulated by temperatures
below 15 or above 45°C.?
3. Chemosensitive pain receptors: Which are stimulated by chemicals like
P-factor, bradykinin, lactic acid, potassium ions ...etc?
All these receptors are naked free nerve endings.
TYPES OF PAIN ACCORDING TO ITS SITE OF ORIGIN
According to the site of its origin, pain is classified into three types:
I. CUTANEOUS PAIN: Which arises from the skin and is transmitted by
cutaneous somatic nerves?

II. DEEP PAIN: Which arises from deep structures like muscles, tendons,
ligaments, periosteum, and is transmitted by deep somatic nerves?
III. VISCERAL PAIN: Which arises from viscera like the heart, the kidney and
the intestine, and is transmitted by the visceral afferent fibers in the autonomic
nerves?
TYPES OF PAIN ACCORDING TO THE SPEED OF ITS CONDUCTION
Pain is conducted by either type III or type IV sensory nerve fibers.
Accordingly, there are two types of pain; fast pain which is conducted by type III
fibers, and slow pain which is conducted by type IV fibers. They differ widely in
their properties and characteristics.
On application of a noxious mechanical stimulus to the skin (e.g. a pinprick on
the foot), one first feels a bright, sharp, localized pain of very short duration (less
than one second). This is the fast pain, which is also called "pricking pain". This
pain is produced by stimulation of the mechanosensitive pain receptors. Shortly
after feeling the pricking pain, one feels a dull, intense, burning, diffuse pain of
longer duration (few sec. to few min.). This is the slow pain, which is also called
"burning pain" or "aching pain". This pain is produced by stimulation of the
chemosensitive pain receptors.
CHARACTERISTICS OF FAST PAIN
1. It is only cutaneous.
2. It is conducted by type-Ill fibers (speed: 6-30 m/s)
3. It is felt immediately after application of the stimulus (0.1 second after
application of the stimulus).
4. It is of very short duration.
5. It is bright and sharp.
6. It is well localized.
7. It is perceived only by the sensory cerebral cortex.

8. It is conducted by the necspinothalamic sensory pathway.
CHARACTERISTICS OF SLOW PAIN
1. It can be cutaneous, deep or visceral.
2. It is transmitted by the slowly conducting type-IV fibers (speed: 0.5-2 m/s).
3. It starts about one second after application of the stimulus.
4. It lasts for a long time.
5. It is dull, aching (or burning).
6. It is diffuse and poorly localized.
7. It can be perceived at the level of the thalamus; the cerebral cortex is not
essential for its perception.
8. It is conducted by the paleospinothalamic sensory pathway.
DISSOCIATION OF FAST AND SLOW PAIN
One type of pain may disappear leaving the other intact, as in the following
conditions:
1. Compression or hypoxia of a nerve abolishes the conduction of fast pain but
leaves slow pain. This is because hypoxia affects the thick type-Ill nerve fibers first
2. Mild local anesthesia (e.g. local low dose of cocaine) abolishes the conduction
of slow pain but leaves fast pain. This is because mild anesthesia affects the thin
type-IV nerve fibers first
I- CUTANEOUS PAIN
Cutaneous pain arises from the skin on application of a noxious stimulus. It is
conducted by somatic cutaneous nerves and is not referred to any other part. It is
usually accompanied by sympathetic reflexes (e.g. increase in heart rate and rise in
arterial blood pressure), but there is no reflex muscular rigidity.
Cutaneous pain could be of the fast "pricking" type or of the slow "burning" type.
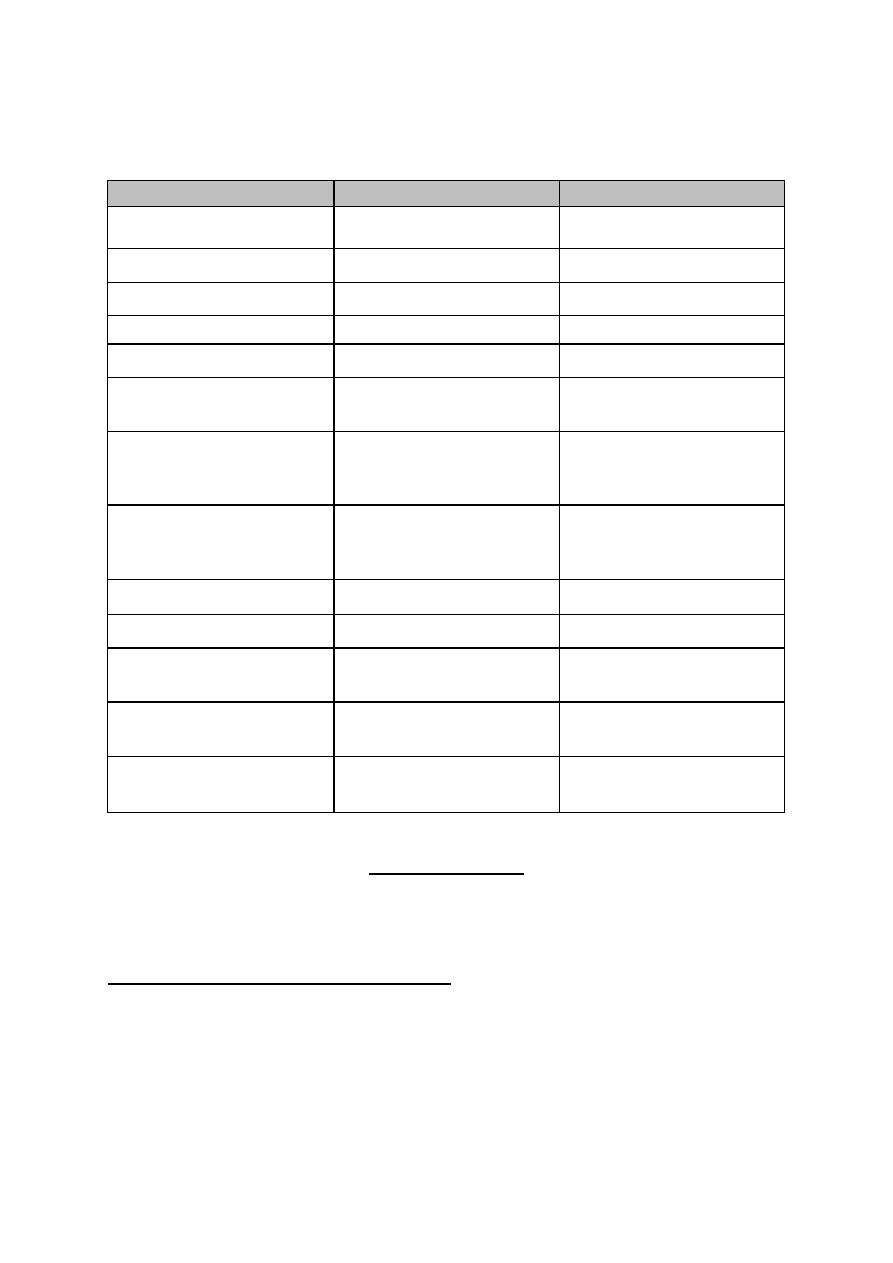
Table 8-1: Differences between, fast and slow pain.
CRITERION
FAST PAIN
SLOW PAIN
1. Site of origin
only cutaneous
cutaneous, deep or
visceral
2. Time of perception
0.1 sec. after the stimulus
1 sec. after the stimulus
3. Characteristics
bright, sharp & localized
dull, intense and diffuse
4. Duration
very short (less than 1
sec.)
longer (few sec. - few
min.)
5. Conducting nerves
type-HI fibers
type-IV fibers
6. Sensory pathway
neospinothalamic
pathway
paieospinothalamic
pathway
7. Associated reflexes
rapid,
protective withdrawal
reflexes
slower
autonomic reflexes (e.g.
increase in heart rate &
ABP)
8. Abolished by
hypoxia or pressure on
the nerve
mild local
anesthesia (low dose
of local cocaine)
9. Center of perception
cerebral cortex
thalamus
10. Relay in dorsal horn
laminae II & V
laminae II & III
11.Relay nuclei in the
thalamus
VPLN
ELN
12.Relay in
reticular
formation
no relay
relay
13.Final destination in
CNS
sensory cerebral cortex
all areas of the cerebral
cortex.
11 - DEEP PAIN
Deep pain is a slow type of pain which arises from deep structures; i.e. muscles,
tendons, ligaments and periosteum.
CHARACTERISTICS OF DEEP PAIN
1. It is dull, aching, diffuse and poorly localized.
2. Usually associated with nausea, sweating and hypotension.
3. Pain from tendons, ligaments and periosteum initiates reflex muscular
spasm in the nearby muscles.

4. It is conducted by the slowly conducting type-IV fibers.
5. It may be referred to the skin of the same dermatome.
CAUSES OF DEEP PAIN
1. Ischemia of skeletal muscles: This leads to the release and accumulation of
P-factor (Lewis pain-producing factor). The nature of this factor is not exactly
known, but it could be bradykinin, histamine, proteolytic enzymes, lactic acid or
potassium ions or a collection of all these substances.
P-factor directly stimulates the chemosensitive pain receptors, but it also
facilitates the mechanosensitive and thermosensitive pain receptors.
Prostaglandins are released from damaged and inflamed tissues. They facilitate
but do not stimulate pain receptors. So they produce tenderness of the affected
area.
INTERMITTENT CLAUDICATIONS (LAMENESS OR LIMPING):
This disease is an example where there is severe ischemic pain in the muscles of the
lower limb. In this condition, there is narrowing of blood vessels of the lower limbs
(following vascular inflammation or severe atherosclerosis). The slightest exercise
leads to accumulation of the P-factor and pain sensation then limping. . During rest", there
is no. pain as there is no accumulation of the P-factor.
2. Muscular spasm (cramp): In tetany or when one loses a lot of salt in sweating
(e.g. with muscular exercise in hot weather or miner's disease) muscle spasm
(cramp) occurs and stimulation of mechanosensitive pain receptors then pain
sensation. Ischemia also contributes to the production of pain in this condition.
3. Bone injury: It is painful because of the chemical and mechanical irritation of
the overlying periosteum. Bone itself is insensitive to pain.
4. Inflammation of joints: It is painful due to chemical and mechanical irritation

of the surrounding tendons and ligaments.
III - VISCERAL PAIN
Visceral pain is a slow type of pain that arises from the viscera (abdominal and
thoracic contents). It is carried by the visceral afferent type-IV slow conducting nerve
fibers. Some regions are insensitive to pain as the liver parenchyma and lung
alveoli.
CHARACTERISTICS OF VISCERAL PAIN
1. It is diffuse, dull or colicky.
2. It is a slow pain, carried by the slowly conducting type-IV fibers.
3. It is poorly localized.
4. It may be referred to the skin or other deep structures.
5. It is associated with:
(a) Autonomic reflexes: Nausea; vomiting, sweating, bradycardia. That is why
visceral pain is said to be "sickening".
(b) Somatic reflexes: Rigidity of the overlying skeletal muscles; e.g. the rigidity
of muscles over the inflamed appendix (guarding rigidity).
(c) Cutaneous hyperalgesia: Increased pain sensibility in the skin of the same
dermatome.
(d) Strong emotional affect: With depression, malaise and bad mood.
NERVES CONDUCTING VISCERAL PAIN
Visceral pain is conducted via afferent visceral fibers in the autonomic nerves (fig.
8-1). Somatic nerves conduct pain from some parts of the viscera.
VISCERAL PAIN CONDUCTED BY SYMPATHETIC NERVES
Afferent fibers that run with sympathetic nerves conduct pain from the thoracic and
abdominal viscera, the urinary bladder, uterus and uterine tubes.
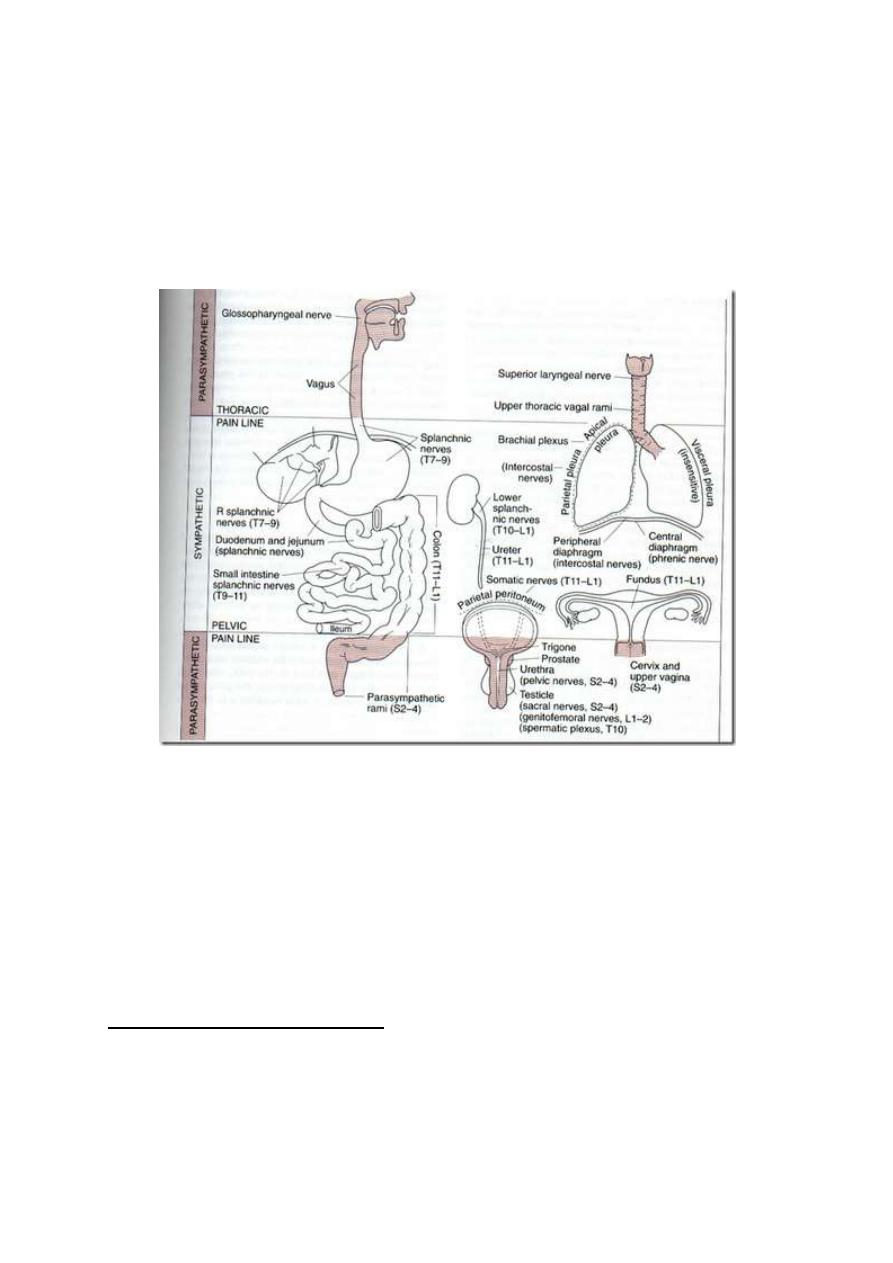
VISCERAL PAIN CONDUCTED BY PARASYMPATHETIC NERVES
• With glossopharyngeal and vagus nerves (Cr. IX and X nerves):
Pain from pharynx, trachea and upper part of the esophagus
• With the pelvic nerve (S 2, 3 & 4): Pain from sigmoid colon,
rectum, the trigone of the bladder and the genital organs.
Figure 8 -1: The areas from which pain is conducted by somatic and by autonomic nerves.
VISCERAL PAIN CONDUCTED BY SOMATIC NERVES
Thoracic intercostal nerves carry pain sensations from parietal pleura, parietal
pericardium and parietal peritoneum. This pain is called "parietal pain". It is
well localized.
CAUSES OF VISCERAL PAIN
1. Ischemia: Ischemic pain is due to accumulation of the P-factor which
stimulates the chemosensitive pain receptors. E.g. anginal pain of myocardial
ischemia.

2. Spasm of a hollow viscus: Spasmodic pain is colicky pain caused mainly by
stimulation of the mechanosensitive pain receptors. Ischemia due to squeezing of
blood vessels may contribute to pain production. E.g. intestinal, renal or biliary
colic.
3. Overdistention of a hollow viscus: Overstretch of the tissues stimulates
mechanosensitive pain receptors. Ischemia may contribute. E.g. overdistention of
the intestine by large volumes of gases.
4. Chemical irritation: Chemical irritants produce pain by stimulating
chemosensitive pain receptors. E.g. Rupture of a peptic ulcer could lead to leak of
the highly acidic gastric juice into the peritoneum leading to severe abdominal
pain. Inflammations produce pain by producing chemical irritants (pain
producing substances).
5. Traction on the mesentery: E.g. by an abdominal tumor. This cause is rare
and of little clinical significance.
REFERRED PAIN
Referred pain is pain felt away from its site of origin.
Examples:
1. Ischemia of the heart causes pain that may be referred and felt in the left
shoulder and the inside of the left arm.
2. Pain from the gall bladder may be referred and felt in the right shoulder.
3. Pain of appendicitis may be referred to the umbilical area.
4. Pain from the ureter may be referred to the testis.
MECHANISM OF REFERRED PAIN (convergence projection mechanism):
Afferent pain-conducting fibers from the viscera converge with afferent
pain-conducting afferent pain-conducting fibers from the skin on one central
neuron of the paleospinothalamic tract.

Figure 5-2: An example of the convergence projection mechanism of referred pain.
In this way, pain impulses from the viscera travel in the same central pathway as
pain impulses from the skin to reach the same final sensory neuron in the brain.
The final sensory neuron projects pain sensation to the skin as the skin is the place
from which it usually receives pain signals (fig. 5-2).
This means that the pain signals from the viscera converge to the same final
sensory neuron as the signals from the skin, the brain projects any pain sensation
from the viscera to the skin. This mechanism is known as "the convergence
projection mechanism".
Accordingly, if pain arises from a certain viscera from which impulses converge
with impulses coming from a certain area of skin, one would feel pain as if it is
coming from the skin not from the viscera.
Reference of pain occurs from viscera to deep structures or the skin and from
deep structures to the skin, but not the opposite way.

DERMATOMAL RULE OF REFERRED PAIN
Visceral or deep pain is referred to the dermatome which has the same nerve
supply as the site of pain. E,g. the heart, the left shoulder and the inside of the left
arm have the same nerve supply. So, pain from the heart is referred to the left
shoulder and the inside of the left arm (fig. 5-2).
THE GATE THEORY OF PAIN TRANSMISSION
The dorsal hom of the spinal gray matter is the site of first relay in the sensory
pathway of pain. At this level, there is a group of inhibitory, enkephalinergic
intemeurons which form the "Pain. Inhibitory Complex, PIC. When
stimulated, these interneuron's block the transmission of pain sensation by
presynaptic inhibition of the primary pain-conducting fibers. At the dorsal horn
also, transmission of pain sensation could be facilitated in cases of secondary
hyperalgesia. In this way, the dorsal horn acts as a "gate" that may pass, facilitate
or block pain transmission. Accordingly, the dorsal horn of the spinal gray matter
is sometimes referred to as the "gate" for pain transmission.
The same "gating" mechanism for pain is found also at the thalamus where pain
signals could be blocked by corticofugal fibers or facilitated by intralaminar
thalamic nuclei. In this way, the thalamus acts as a secondary gate for pain
transmission.

Physiology
Dr. Basim Mohamad Alwan Lecture (6)
INTERINSIC MODIFICATION
O
F PAIN SENSIBILITY
Intrinsic mechanisms can modify pain sensation leading to exaggerated or
suppressed pain sensibility.
I. EXAGGERATED PAIN SENSIBILITY
1. CUTANEOUS HYPERALGESIA
This is a pathological condition where pain sensibility from the skin is exaggerated.
It usually follows a skin injury or inflammation. There are two types of cutaneous
hyperalgesia:
A. PRIMARY HYPERALGESIA
In this case, the threshold of pain sensation from the affected area is lowered. A
stimulus which normally produces mild pain causes severe, prolonged pain.
Primary hyperalgesia occurs in conditions like sunburns. It is restricted to the
affected area of skin and the area of hyperemia around it (the area of the spreading
flare). This type of hyperalgesia is caused by facilitation of pain receptors by
substances released from the damaged or inflamed tissues, e.g. histamine,
bradykinins, substance-P, Prostaglandins ...etc.
ALLODYNIA
is a severe type of primary hyperalgesia in which a gentle stimulus like
a breeze or the touch of clothes produces severe pain.
B. SECONDARY HYPERALGESIA (HYPERPATHIA)
In this case, the threshold of pain is increased, but when the threshold is reached it
produces severe burning pain. It occurs in a normal skin area that extends beyond the
spreading flare of the injured skin. Secondary hyperalgesia is produced by
convergence-facilitation mechanism (fig. 6-1).
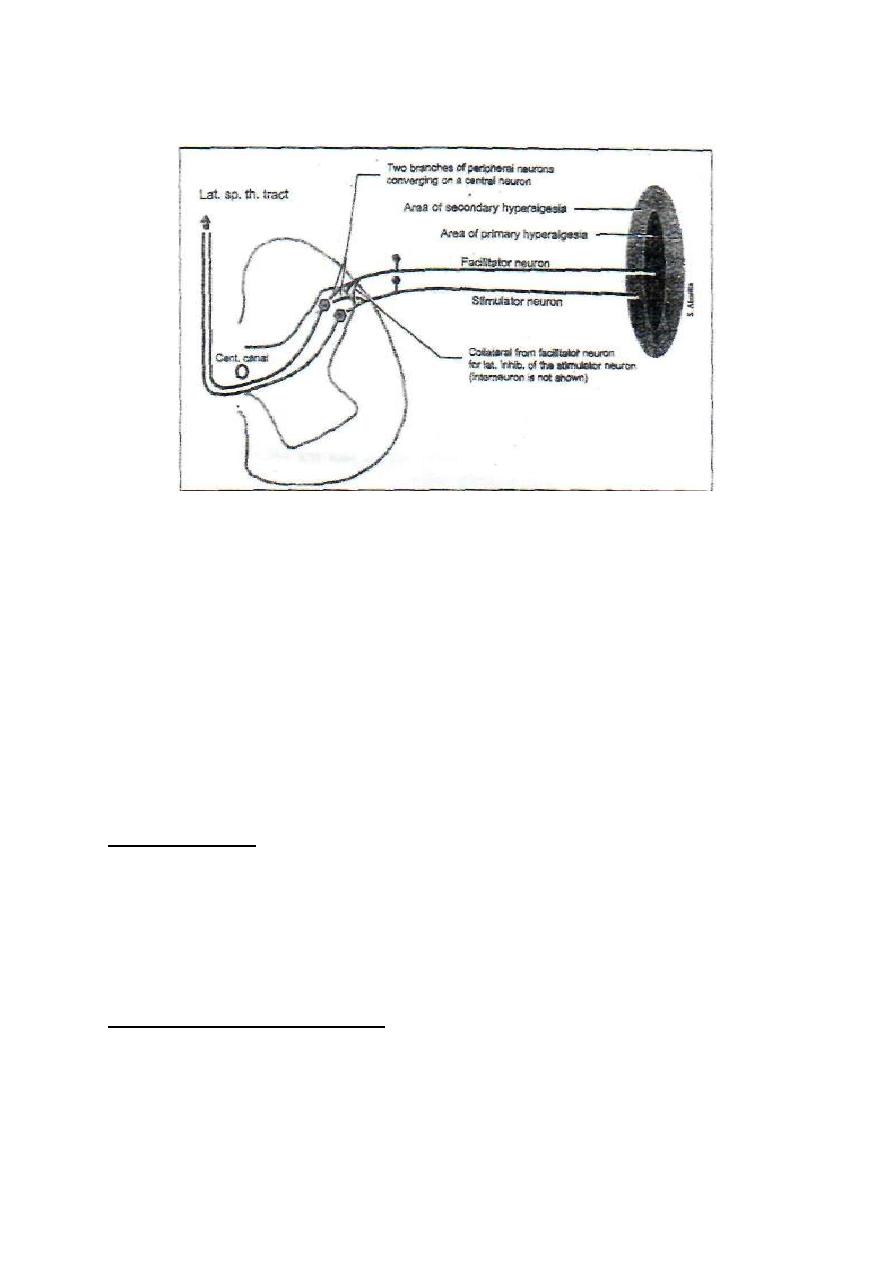
Figure 6-1: The convergence facilitation mechanism of secondary hyperalgesia.
Impulses from the injured area facilitate a central neuron. Impulses from the
hyperpathic area converge on the same central neuron. The convergence on a
central facilitated neuron explains the exaggerated pain sensibility. The
facilitator neuron which arises from the area of primary hyperalgesia exerts
lateral inhibition on the stimulator neuron which arises from the hyperpathic
area. This explains why the threshold of pain is increased in the hyperpathic area.
2. CAUSALGIA
Causalgia is a pathological condition in which there is hyperalgesia, allodynia
and spontaneous burning pain sensation long after a seemingly trivial skin injury.
It occurs if the skin injury was associated with nerve injury. The skin in the
affected area becomes thin and shiny with increased hair growth.
THE MECHANISM OF
CAUSALGIA:
Nerve injury leads to sprouting of noradrenergic sympathetic nerve fibers into the
sensory nerve track up to the dorsal root ganglia of the sensory nerves of the
affected area. The dorsal root ganglia cells can be stimulated by sympathetic

activity. This stimulation may facilitate the sensory neuron leading to hyperalgesia
or allodynia, or it may excite it to discharge pain signals to the CNS leading to
spontaneous pain sensation.
3. THE THALAMIC SYNDROME
Thalamic syndrome is caused by thrombosis of the thalamogeniculate artery, a
branch of the posterior cerebral artery. At first, all sensations are lost on the
opposite side. Few weeks later, pain sensibility is regained. A noxious stimulus
produces very severe, stabbing and extremely unpleasant pain. This is due to the
facilitation of the medial and laminar nuclei of the thalamus which do not degenerate
in this syndrome. They potentiate pain conducted by the reticular activating system of
the brainstem.
II. SUPPRESSED PAIN SENSIBILITY
ANALGESIA
Analgesia means suppression of pain sensation. There is an analgesia system in
the body which can suppress pain sensibility by activating the spinal pain
inhibitory complex. The analgesia system of the body is divided into peripheral
and central analgesia systems.
THE PERIPHERAL ANALGESIA SYSTEM
The peripheral anesthesia system consists of the type-II sensory nerve fibers which
conduct mechanoreceptive sensations.
Immediately after they enter into the spinal cord, type-H fibers divide into medial
and lateral branches. The medial branch ascends to form the gracile and cuneate
tracts, and the lateral branch stimulates the PIC to inhibit pain transmission. In
this way, pain sensation can be suppressed by scratching or rubbing of the skin.
Counterirritants which are applied to the skin to suppress pain (mustard plaster
or liniments) work on this principle. They stimulate the cutaneous receptors

which send impulses through type-II fibers that stimulate the PIC. This is also the
basis for acupuncture.
On the same principle, electrodes surgically implanted in the dorsal column
activate the PIC and suppress pain sensibility. They are sometimes used for
treatment of intractable pain.
THE CENTRAL ANALGESIA SYSTEM
This system consists of five components (fig. 6-2)
1. THE CEREBRAL CORTEX
Many areas of the cerebral cortex, especially the limbic association area, project
corticofugal fibers to:
a. The thalamus; these fibers block pain signals at the level of the thalamus (the
second gate for pain transmission).
b. The periaquiductal gray matter of the mesencephalon.
The corticofugal fibers are B-endorphinergic.
2. THE HYPOTHALAMUS
B-endorphinergic fibers project from the periventricular nuclei and the medial
forebrain bundle of the hypothalamus to the periaquiductal gray matter of the
mesencephalon.
3.
THE PERIAQUEDUCTAL GRAY MATTER OF THE MIDBRAIN
Neurons from this area project enkephalinergic fibers to the reticular formation of the
brainstem and fee raphe magnus nucleus.
4. THE RETICULAR FORMATION OF THE BRAINSTEM
This structure sends serotonergic descending fibers in the lateral reticulospinal
tract to activate the spinal PIC. It also projects fibers to the raphe magnus nucleus.
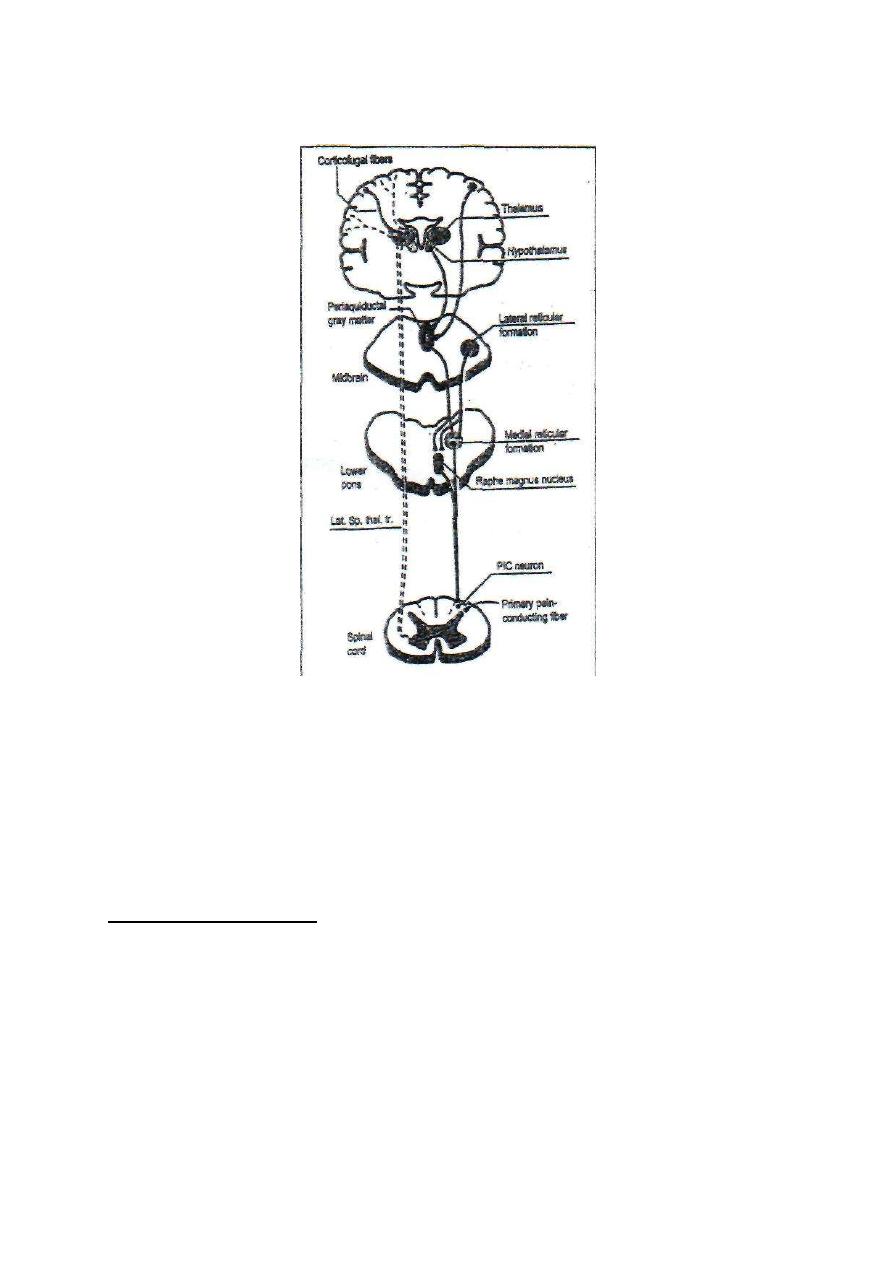
Fig.6-2 the central analgesia system
5. THE RAPHE MAGNUS NUCLEUS
The raphe magnus nucleus is found in the lower pons and upper medulla.
Serotonergic nerve fibers descend from this nucleus down in the lateral reticulospinal
tract to activate the spinal PIC.
STRESS ANALGESIA
Stress analgesia is the suppression of pain sensation during stressful during stressful
conditions (fight or flight). In these conditions, pain impulses are blocked at two
levels:
A. At the first gate of pain transmission (the dorsal horn of the spinal gray
matter). The hypothalamus and other parts of the central analgesia system
activate the spinal PIC which blocks the transmission of pain signals at the

dorsal horn.
B. At the second gate of pain transmission (the thalamus). Corticofugal fibers
to the thalamus block by presynaptic inhibition the transmission of pain signals in
the thalamus before they reach the cerebral cortex.
THE NERVOUS PATHWAYS OF SOMATIC SENSATIONS
A tract is a bundle of nerve fibers in the spinal cord with common origin, common
destination and a common function. An ascending tract is a sensory tract which
conducts sensory signals up to the higher brain centers. A descending tract is a
motor tract which conducts motor signals from the higher brain centers down to the
motor neurons in the motor neuron pool.
The nervous pathway of a sensation starts from its sensory receptors up to the
final sensory neurons in the brain. It involves 2 - 4 orders of neurons. The first
order neurons of all sensory pathways are the afferent sensory nerves which have
their cell bodies in the dorsal root ganglia or equivalent ganglia of the cranial
nerves. An ascending tract is the part of the sensory pathway which is found in
the spinal cord.
THE VENTRAL SPINOTHALAMIC PATHWAY
The ventral spinothalamic pathway (fig. 6-3) is the sensory pathway for crude
touch, tickle and itch sensations. It has three orders of neurons:
FIRST ORDER NEURONS:
are the dorsal root neurons. Their afferent fibers (type III and
IV) enter the spinal cord in the dorsal root of the spinal nerve then go up or down for
a few segments in the Lissaur s tract and terminate in laminae II, III and IV of
the dorsal horn of the spinal gray matter.
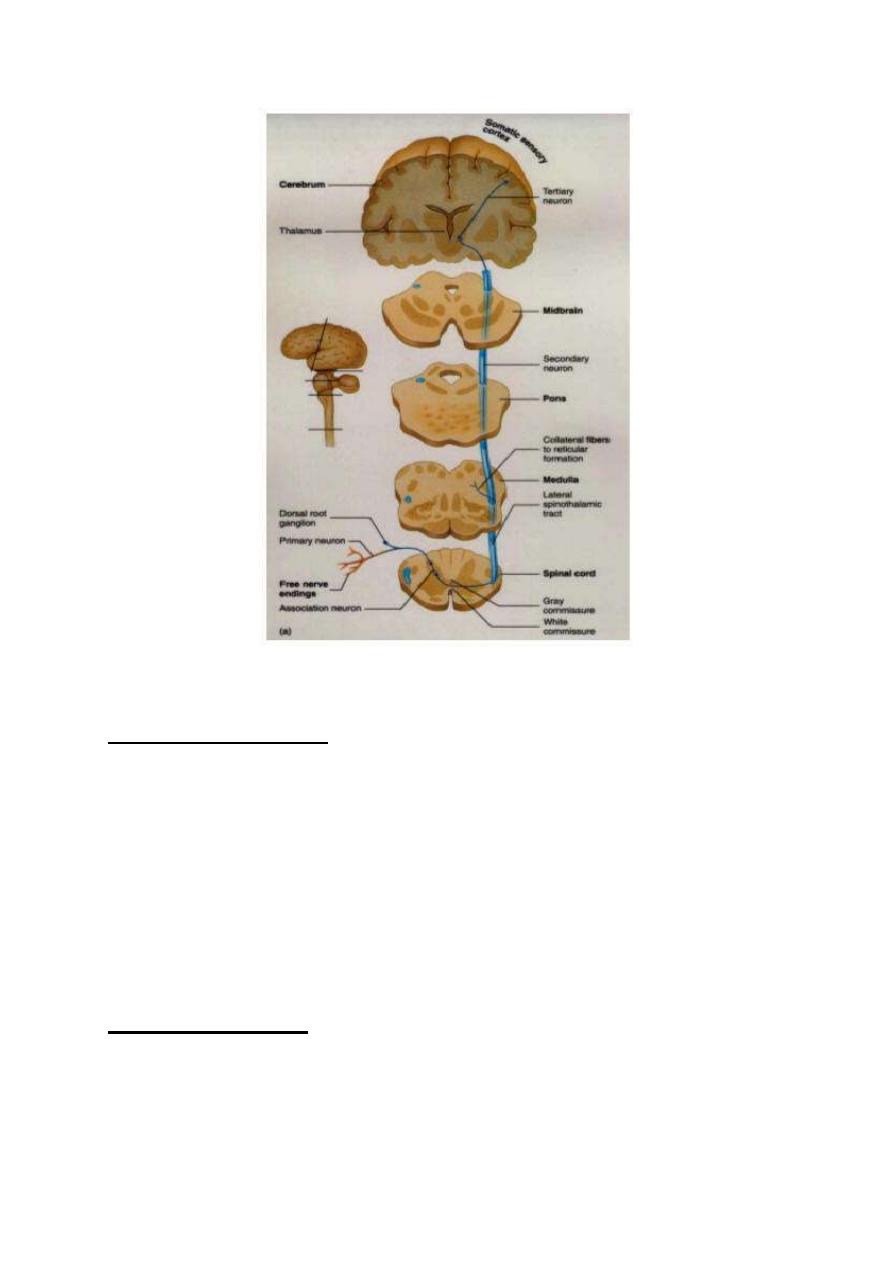
Fig. 6-3 ventral spinothalamic tract
SECOND ORDER NEURONS:
are neurons in laminae II, III and IV of the dorsal horn.
Their fibers cross to the opposite side in front of the central canal then ascend in
the ventral column of the spinal white matter as the ventral spinothalamic tract up
to the brainstem and in the brainstem, they join the lateral spinothalamic tract to
form the spinal lemniscus (nervous tract made by union of more than one
tract) which ascend up to terminate in the ventral posterolateral nucleus of the
thalamus (VPLNT).
THIRD ORDER NEURONS
: are neurons of the VPLNT. Their fibers project through
the central thalamic radiations to the final sensory neurons in the sensory cerebral
cortex.
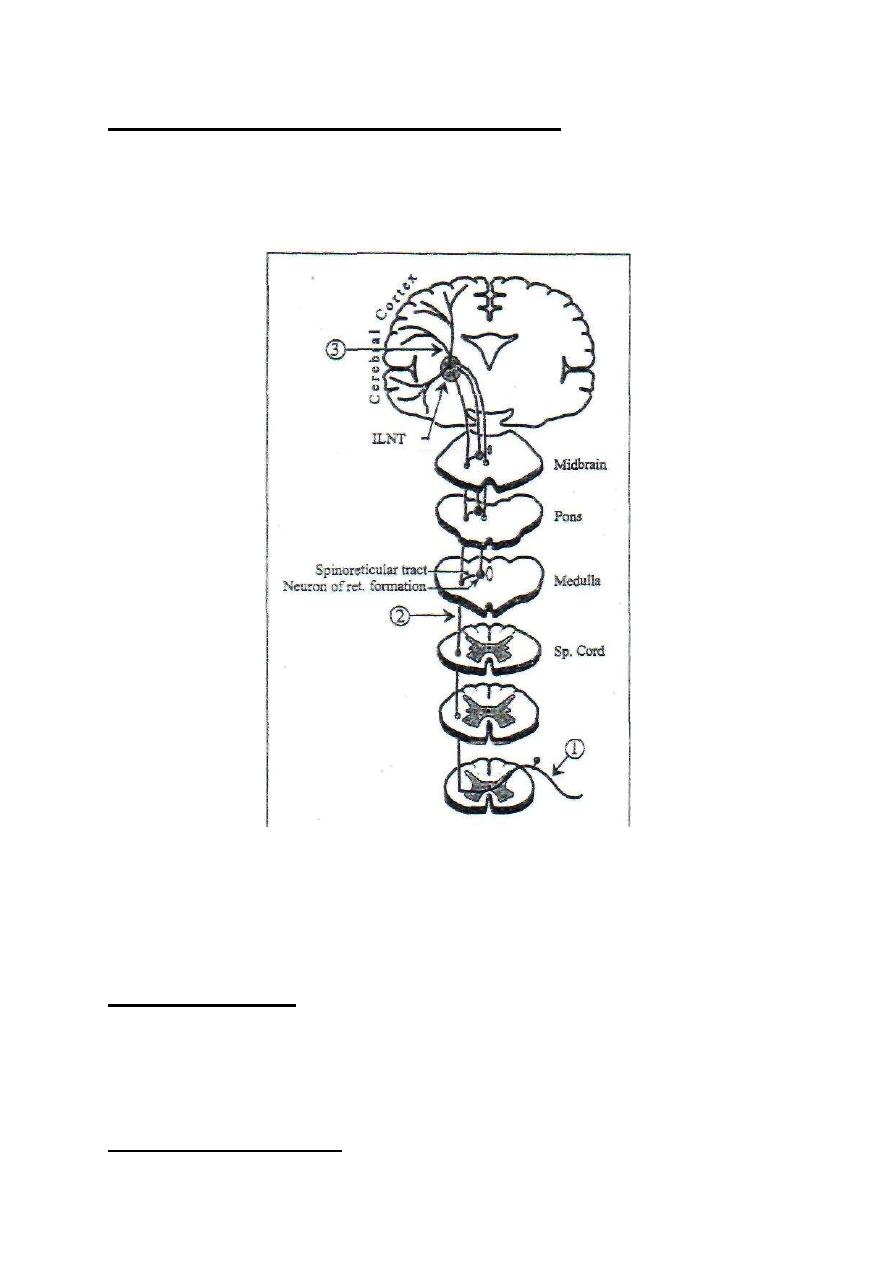
THE LATERAL SPINOTHALAMIC PATHWAYS
There are two lateral spinothalamic pathways; the paleospinothalamic and the
neospinothalamic pathways.
1. THE PALEOSPINOTHALAMIC PATHWAY
Fig. 6-4 the paleospinothalamic pathway
The paleospinothalamic pathway (fig. 6-4 is the sensory pathway for slow pain,
and temperature sensations.
FIRST ORDER NEURONS:
are the dorsal root neurons. Their afferent fibers (type IV)
enter the spinal cord in the dorsal root of the spinal nerve then go up or down for a
few segments in the Lissaur tract and terminate in the substantia gelatinosa of
Rolandi (laminae II, III) of the dorsal horn of the spinal gray matter.
SECOND ORDER NEURONS:
are neurons in the substantia gelatenosa of Rolandi.

Their fibers cross in front of the central canal to the opposite side. Fibers ascend in
the anterolateral column of the spinal white matter as the lateral spinothalamic
tract. In the brainstem, the lateral spinothalamic tract. In the brainstem the
lateral spinothalamic tract joins the ventral spinothalamic tract to form the spinal
lemniscus. Fibers terminate in the nonspecific intralaminar nuclei of the
thalamus (ILNT).
During their course in the brainstem, some fibers deviate and make a separate tract
called "the spinoreticular tract". The fibers of this tract terminate in the reticular
formation of the brainstem. Multiple, short fiber neurons conduct the signals from
the reticular formation onto the intralaminar nuclei of the thalamus (ILNT).
THIRD ORDER NEURONS
: are neurons of the ILNT. Their fibers project through the
anterior, central and posterior thalamic radiations to the final sensory neurons in all
parts of the cerebral cortex.
2. THE NEOSPINOTHALAMlC PATHWAY
The neospinothalamic pathway (fig. 6-5) is the sensory pathway for fast pain.
FIRST ORDER NEURONS:
are the dorsal root neurons. Their afferent fibers (type
III) enter the spinal cord in the dorsal root of the spinal nerve and go up or down for
a few segments in the Lissaur tract then terminate in laminae II and V of the dorsal
horn of the spinal gray matter.
SECOND ORDER NEURONS:
are neurons in laminae, II and V. Their fibers cross in
front of the central canal to the opposite side, and then ascend in the anterolateral
column of the spinal white matter as the lateral spinothalamic tract. In the
brainstem, the lateral spinothalamic tract joins the ventral spinothalamic tract to
form the spinal lemniscus. The fibers of the neospinothalamic tract terminate in
the ventral posterolateral nuclei of the thalamus (VPLNT).
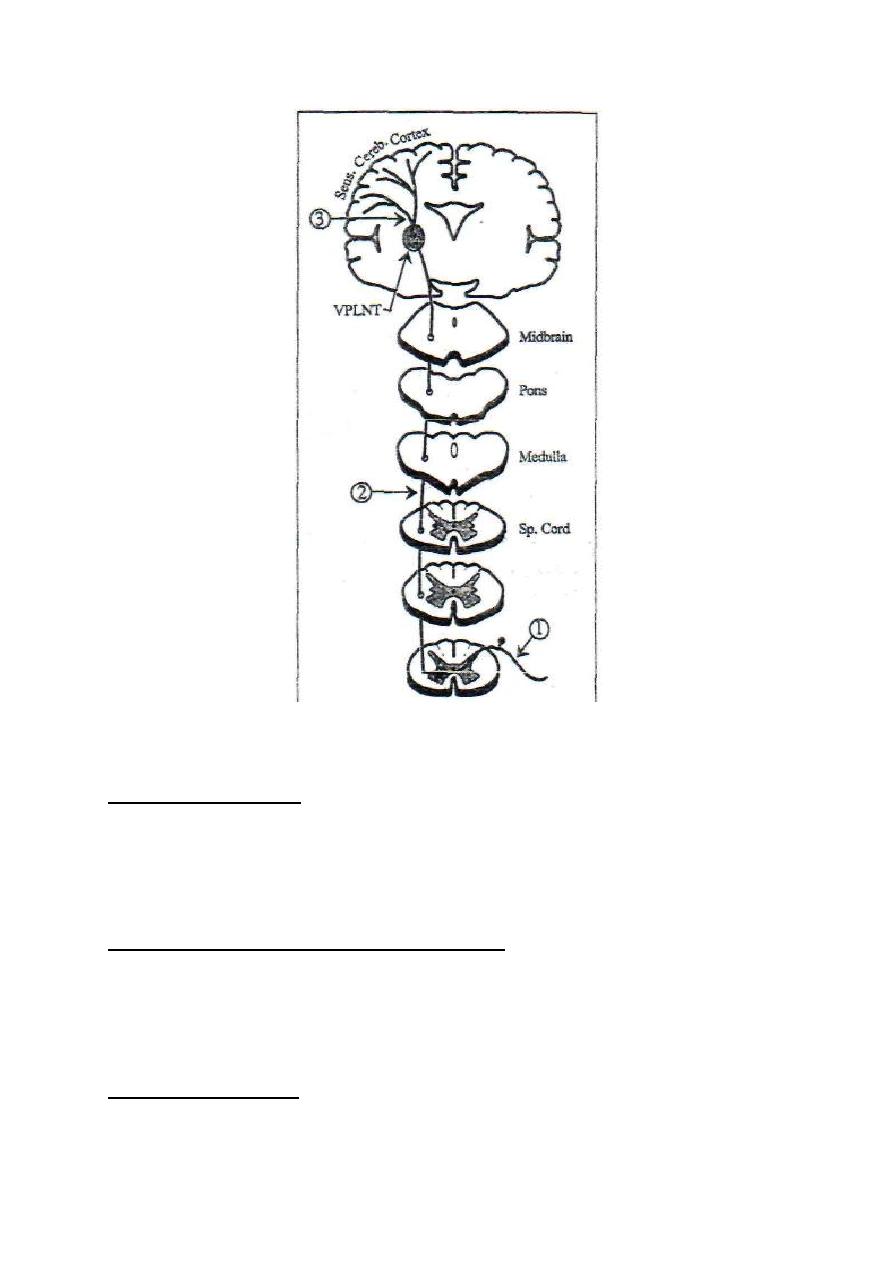
Fig. 6-5 the neospinothalamic pathway
THIRD ORDER NEURONS:
are the neurons of the VPLNT. Their fibers project through
the central thalamic radiations to the final sensory neurons in the sensory cerebral
cortex.
THE DORSAL COLUMN PATHWAY
The dorsal column pathway (also called the graeile and cuneate pathway) (fig. 6-6) is
the sensory pathway for fine touch, fine pressure, vibration, stereognosis, muscle
tension and proprioceptive sensations.
FIRST ORDER NEURONS:
are the dorsal root neurons. Their afferent fibers (type II)
enter the spinal cord in the dorsal root of the spinal nerve then branch into medial
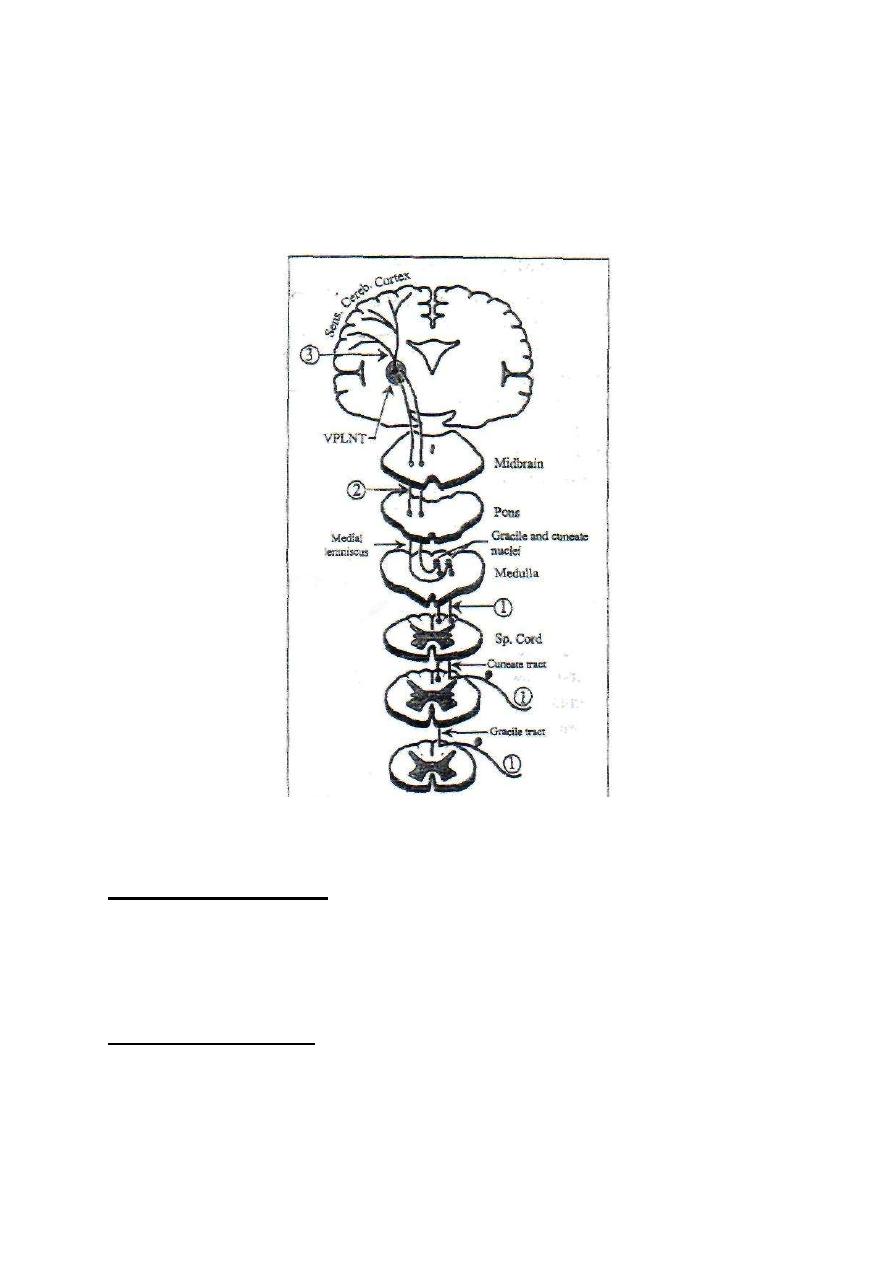
and lateral branches. The medial branches ascend without relay up in the ipsilateral
dorsal column of the spinal white matter where they are called "the dorsal column
tracts" or "the gracile and cuneate tracts". They terminate in the dorsal
column nuclei (the gracile and cuneate nuclei) in the medulla.
Fig. 6-6 the dorsal column pathway
SECOND ORDER NEURONS:
are the neurons of the dorsal column nuclei in the
medulla. Their fibers cross to the opposite side in the sensory decussation and
ascend in the brainstem as the medial lemniscus. They terminate in the ventral
posterolateral nucleus of the thalamus (VPLNT). .
THIRD ORDER NEURONS:
are those of the VPLNT. Their fibers project through the
central thalamic radiations to the final sensory neurons in the sensory cortex.
* The gracile is the medial tract. It is formed in the lower part of the spinal cord and carries
sensations from the lower part of the body. The cuneate is the lateral tract. It is formed in

the upper part of the spinal cord at the level of the 6th thoracic spinal segment, and carries
sensations from the upper part of the body.
THE SPINOCERVICAL PATHWAY
The spinocervical pathway (fig. 6-7) is an accessory pathway for the dorsal column
pathway. It conducts impulses at a faster rate. So, impulses in this pathway reach the
cerebral cortex before those conducted by the dorsal column pathway.
FIRST ORDER NEURONS:
are the dorsal root neurons. Their afferent fibers (type I and
II) enter the spinal cord in the dorsal root of the spinal nerve and terminate in
lamina IV" of the dorsal horn of the spinal gray matter.
SECOND ORDER NEURONS:
are neurons in lamina IV. Their fibers ascend in the
ipsilateral posterolateral column of the spinal white matter as the spinocervical
tract. The fibers terminate in the lateral cervical nucleus
(a longitudinal cell column
lateral to the tip of the dorsal horn of the upper 2-3 cervical segments)
of the same side.
THIRD ORDER NEURONS:
are those of the lateral cervical nucleus. Their fibers
cross to the opposite side and ascend in the brainstem as part of the medial
lemniscus to terminate in the VPLNT.
FOURTH ORDER NEURONS
: are neurons of the VPLNT. Their fibers project
through the central thalamic radiations to the final sensory neurons in the sensory
cerebral cortex.
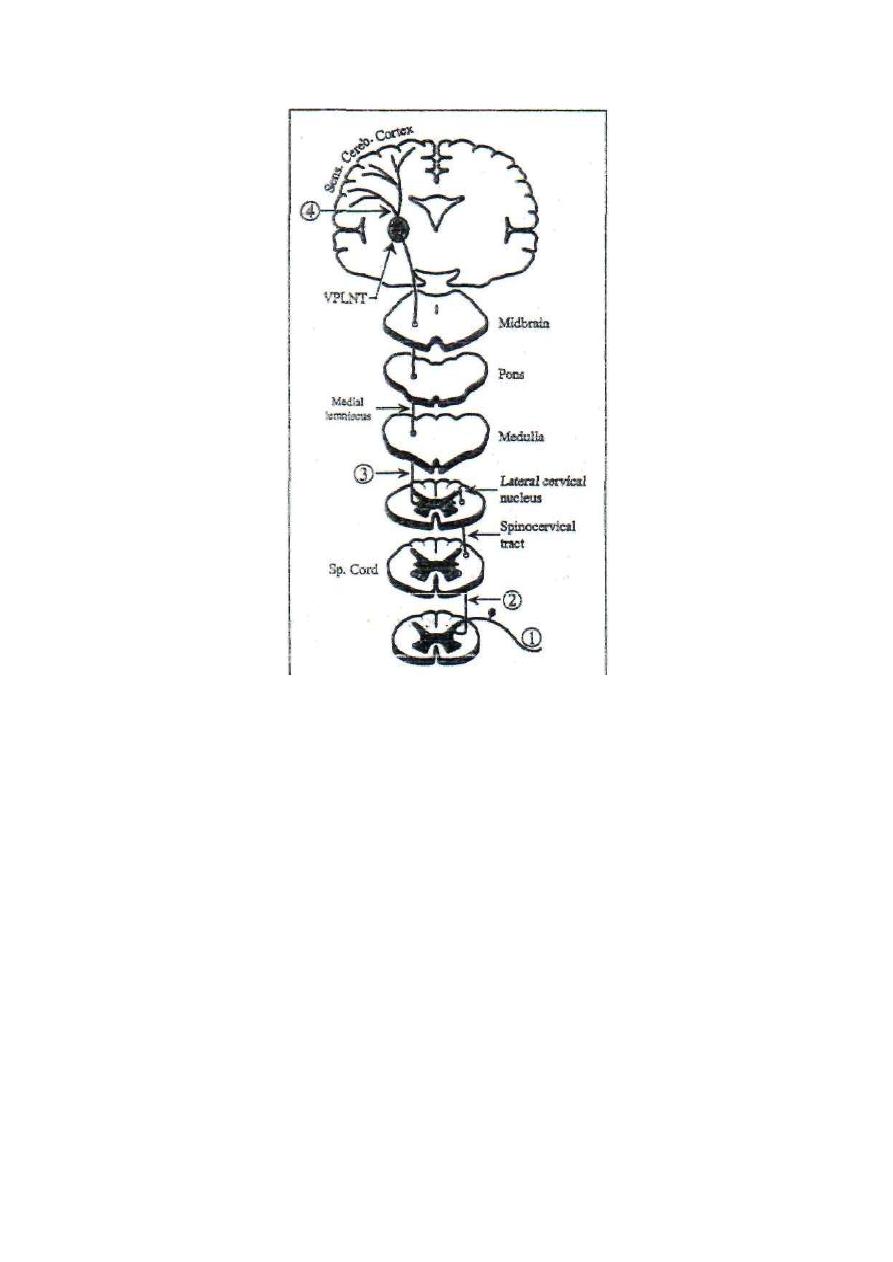
Fig. 6-7 the spinocervical pathway

Physiology
Dr. Basim Mohamad Alwan Lecture (7)
THE SOMATOSENSORY CONDUCTING SYSTEMS OF THE
SPINAL CORD
The ascending somatosensory tracts in the spinal cord are classified into
two conducting systems, i.e. the anterolateral and the dorsal
column-lemniscal systems. Each system has its own features and
characteristics.
THE ANTEROLATERAL SYSTEM
The anterolateral system has two components; the ventral and the lateral
spinothalamic tracts. The fibers of the first order neurons which serve
this system are the slow, thin, myelinated type-III and the
nonmyelinated type-IV fibers.
CHARACTERISTICS OF THE ANTEROLATERAL SYSTEM
1. It is a slowly conducting system made up of thin type-III and IV
fibers.
2. It has moderate degree of somatotopic lamination. This allows only
moderate degree of localization and discrimination of its sensations.
3. The intensity discrimination of its sensations is poor. Sensations of this
system can be identified in only 10-20 grades of intensity.
4. It cannot transmit rapidly repetitive signals.
5. It conducts sensations from the contralateral side of the body.

THE DORSAL COLUMN-LEMNISCAL SYSTEM
The dorsal column-lemniscal system has three components; the dorsal
column tracts (gracile and cuneate), the spinocervical tract, and the medial
lemniscus. The fibers of the first order neurons which serve this system are
the rapidly conducting, thick, myelinated, type-I and II fibers.
CHARACTERISTICS OF THE DORSAL COLUMN LEMNlSCAL SYSTEM
1. It is a rapidly conducting system made up of the thick type-I and II
fibers.
2. It has a high degree of somatotopic lamination; i.e. it is arranged in
many laminae, each lamina serves a specific topic on the body
surface. This allows a high degree of localization and discrimination of
its sensations.
3. The intensity discrimination of its sensations is high. Most sensations
in this system can be identified in up to 100 grades (100 separate
degrees of intensities).
4. It can conduct rapidly repetitive signals. This enables this system to
conduct vibration sense.
5. It conducts sensations from the ipsilateral side of the body.
THE THALAMUS
The two thalami are large ovoid masses of gray matter situated at the
lateral walls of the third ventricle, one on each side. They are
interconnected by a short communicating bar of white matter (massa
intermedia) which traverses the third ventricle.
All the nervous signals which go to the cerebral cortex pass first
through and relay in the thalamus. That is why the thalamus is
sometimes called the "secretary of the cerebral cortex ".
ANATOMICAL DIVISIONS OF THE THALAMUS
In each thalamus there are 5 groups of nuclei:
1. Anterior group: This is connected to the cingulate gyrus and
forms part of the limbic circuit concerned with recent memory.
2. Medial group: This includes the dorsomedial nucleus, the
intralaminar and middle nuclei.
3. Lateral group: This includes the dorsolateral and posterolateral
nuclei.

4. Ventral group: Which includes?
(a) The ventroanterior (VA) nucleus.
(b) The ventrolateral (VL) nucleus.
(c) The ventroposterior (VP) complex (ventrobasal complex), which
includes:
(i) Ventral posterolateral (VPL) nucleus.
(ii) Ventral posteromedial (VPM) nucleus.
5. Posterior group (metathalamus): Which includes the medial and
lateral geniculate bodies.
Various nuclear groups of the thalamus are freely interconnected by
short intermediary neurons.
FUNCTIONAL DIVISIONS OF THE THALAMUS
Functionally, the thalamic nuclei could be classified into four
categories:
[I] Specific projection nuclei (cortical relay nuclei):
a. Ventral posterolateral nucleus (VPL) nucleus, which is the site of
relay of the somatic sensory pathways from the trunk and limbs
b. Ventral posteromedial nucleus (VPM) nucleus, which is the site
of relay of the trigeminal sensory pathway from the head.
c. Medial geniculate body (MGB); site of relay of the auditory
pathway.
d. Lateral geniculate body (LGB): site of relay of the visual pathway.
e. Dorsomedial nucleus; site of relay for the olfactory pathway.

AFFERENT CONNECTIONS:
From the medial, spinal and trigeminal
lemnisci, optic tract, hearing and olfactory pathways.
EFFERENT CONNECTIONS:
Specific thalamic projection system to
specific points in the somatic audio and visual sensory areas of the
cerebral cortex.
[II] Nonspecific projection nuclei:
a. Intralaminar and midline nuclei.
b. Ventroanterior nucleus.
c. Anterior nuclei.
AFFERENT CONNECTIONS:
From the ascending reticular activating
system (ARAS), and the paleospinothalamic tracts.
EFFERENT CONNECTIONS:
Nonspecific thalamic projection fibers to
all parts of the cerebral cortex. The anterior nuclei are connected to the
hypothalamus and limbic system.
[III] Association nuclei:
a. Dorsolateral nucleus.
b. Posterolateral nucleus.
AFFERENT CONNECTIONS:
From other thalamic nuclei
EFFERENT CONNECTIONS:
To prefrontal, parietal and occipital
cortical association areas. No subcortical connections.
[IV] Motor nuclei:
The most important of these is the ventrolateral (VL) nucleus.
AFFERENT
CONNECTIONS: From the basal ganglia and cerebellum.
EFFERENT CONNECTIONS:
The motor cortex.

FUNCTIONS OF THE THALAMUS
1. It is a relay station for all the sensory pathways in their way to the
cerebral cortex. Sensory signals are processed in the thalamus before
reaching the cortex (thalamus is the secretary of the cortex).
2. It acts as a final sensory center for conscious perception of some
sensations: i.e. slow pain, high or low grades of temperature and crude
touch.
3. Facilitation of the cerebral cortex, raising its excitability up to the
level necessary to do all cerebral functions. Without the thalamus, the
cerebral cortical functions are markedly depressed. The cerebral cortex
and thalamus are sometimes considered as one unit called "the
thalamocortical system".
4. Identification of the stimulus affect (pleasant or unpleasant), and
controlling the emotional and motor reactions to it. This is done in
collaboration with the cerebral cortex, the limbic system and
hypothalamus
5. It is part of the limbic circuit which is concerned with recent memory
and memory search.
6. It is part of the caudate and putamen circuits which control the
motor activity
THE THALAMIC SYNDROME (The thalamic hypersthetic anesthesia)
The most common cause of this syndrome is thrombosis of the
thalamogeniculate artery which is a branch of the posterior cerebral
artery. This leads to degeneration of the posterior and ventral parts of the

thalamus. The manifestations which appear on the contralateral side of
the lesion include:
SENSORY EFFECTS
First, there is complete hemi anesthesia at the onset of the disease. Few
weeks later protopathic sensation (primitive sensations which
include crude touch, pain and high or low temperatures) are
recovered. The threshold of pain is elevated but gives very unpleasant,
exaggerated central effect. Epicritic sensations (fine sensations, which
include fine touch, proprioceptive sensations and intermediate
grades of temperature) are permanently and irreversibly lost.
MOTOR EFFECTS
There is hemiparesis (weakness of the muscles), hemiataxia and
choreoathetoid movements.

SENSORY FUNCTIONS OF THE CEREBRAL CORTEX
The cerebral cortex is the highest center for conscious perception of
fine sensations. Crude sensations such as pain, temperature and crude
touch can be perceived at a lower level in the thalamus. Somatic
sensations are perceived in the parietal lobe, visual sensations are
perceived in the occipital lobe and auditory sensations in the temporal
lobe.
There are three somatic sensory areas in the cerebral cortex; the
primary somatic sensory area (SI), the secondary somatic sensory
area (SII) and the somatic sensory association area.
THE PRIMARY SOMATIC SENSORY AREA (SI):
LOCATION;
In the postcentral gyrus of the parietal lobe (Bro dmann
areas 3, 1 and 2) (fig. 7-1). It receives projection fibers from the
ventrobasal complex of the thalamus (VPLNT and VPMNT). In this
area, there are the final sensory neurons of the somatic sensory
pathways of fine sensations.
BODY REPRESENTATION
: It is crossed and inverted representation
(fig. 7-2). The upper half of the face is bilaterally represented.
Crossed representation means that each half of the body is represented on
the contralateral cerebral hemisphere.
Inverted representation means that upper parts of the body are represented
in the lower part of the cortex and lower parts of the body in upper part of the
cortex.
Figure 7-1: Map of Brodmann areas of the cerebral cortex. It is based on
histological typing of cortical cells (first published in 1909).
The area of representation of each part is proportional to the number of
receptors in this part, not to its size, e.g. the lips and thumb are
represented by relatively large areas, whilst the trunk is represented by
relatively small area.
Figure 7 - 2: Body representation in the postcentral sensory gyrus

Cells in the sensory cortex are arranged in columns projecting inwards
from the surface. Points in the cortex that represent a certain area of the
body contain several columns each column is specialized in perception
of a specific sensory modality. So, modality representation is
included within the topographic representation.
Topographic areas of representation are not permanent or unchan-
geable, but can be changed and modified. E.g., if a finger is amputated,
the cortical representation of the neighboring fingers creep into the area
of the amputated finger.
FUNCTIONS:
The primary somatic sensory area is essential for the
perception of:
1. Fine touch sensation, i.e. tactile localization and discrirnination.
2. Localization of pain and temperature sensations, SI is not essential
for the perception of pain and temperature sensations, it is only needed for
their accurate localization.
3. Intensity discrimination of different stimuli.
4. Texture of material.
5. Proprioception; static and dynamic.
SI area project connection fibers to the secondary somatic sensory area (SII)
and the somatic sensory association area for interpreting the meaning and
significance of the sensory information; E.g. stereognosis.
THE SECONDARY SOMATIC SENSORY AREA (SII)
LOCATION:
In the supramarginal gyrus (Brodmann area 40), behind
mellower part of SI.

BODY
REPRESENTATION:
Bilateral representation with poor
topographic representation. The head area is generally in the anterior part
and the leg area in the posterior part.
SII receives connection fibers which convey input signals from SI, the
visual and auditory cortical sensory areas, and the thalamic nuclei on both
sides of the body.
FUNCTIONS:
SII is a potentiator of SI. It cannot work independent of SI.
SI, however, can carry out its functions without SII. So, SI can work
without SII, but the opposite is not true.
SOMATIC SENSORY ASSOCIATION AREA
(SOMATIC INTERPRETATIVE AREA)
LOCATION:
In the posterior parietal cortex (Brodmann areas 5 and 7),
behind SI and above SII.
CONNECTIONS:
It receives sensory signals from SI, SII, and the thalamus.
FUNCTIONS:
1. It combines all sensory signals to give meaning to the complex
sensory input.
2. Stereognosis. This area is the center of stereognosis.
3. Spatial orientation of the body with its surroundings. The person
recognizes the position of each part of the body relative to other
parts and to the surrounding objects.
4. Memory. This area is the memory store of previous sensory
experience. Stimulation of this area produces sensory hallucinations.
EFFECT OF
LESION
: A lesion in the somatic sensory association area

results in:
1. Astereognosis, i.e. inability to identify objects by their touch, shape,
weight and texture.
L .Autotopagnosia, i.e. loss of recognition of part or whole
contralateral side of the body. The patient does not acknowledge the
existence of the affected part and fails to include it in planning of
voluntary movement. E.g. he does not swing the arm of the affected side
during walking, he puts clothes on one side of the body and ignores the
other or he shaves only one side of his beard.
3. Impaired memory and decreased intelligence.

CELLS AND LAMINAE OF THE CEREBRAL CORTEX
The cerebral cortex is arranged in six distinct laminae according to the
histological architecture of each laminae (fig. 7-3). These laminae are
labeled from outside inwards as lamina I to lamina VI. There are
generally two types of cells in the cerebral cortex (fig. 7-4).
1. The pyramidal cells; which have a pyramidal shape and are found
in layers II, III, V and VI. The special feature of these cells is that their
axons leave the cortex. These axons terminate in one of the following
destinations:
a. Association fibers: which originate from lamina II and terminate
in other cortical areas on the sat
b. Commissural fibers; which originate from lamina III and
terminate in cortical areas on the opposite side.
c. Pyramidal tracts; which originate from lamina V and terminate in
motor nuclei in the brainstem and spinal cord.
d. Cortical projection fibers; which originate from lamina VI and
terminate in subcortical structures; e.g. corticothalamic projections.
2. The stellate cells: which have a star-like shape and are found mainly
in layer IV? Their axons terminate within the cortex. They are the final
cortical sensory neurons which are responsible for conscious
perception of different sensations.
Figure 7-3: The types of cells in different layers of the cerebral cortex and
their connection.
FUNCTIONS OF THE DIFFERENT CORTICAL LAMINAE
Lamina I: It consists of interconnecting nerve fibers which connect
different areas of the cortex. These fibers arise from all other laminae in
the cortex. It also contains fibers from the nonspecific thalamic
nuclei.
Lamina II: Contains pyramidal cells which send association fibers to
other areas of the cortex on the same side.

Lamina III: Contains pyramidal cells which send commissural fibers to
the cortex on the opposite side.
Lamina IV: Consists of stellate cells which receive input fibers from the
specific sensory nuclei of the thalamus. These cells are the final sensory
neurons which are responsible for the conscious perception of different
sensations.
Lamina V: Consists of pyramidal cells whose axons descend as the
motor pyramidal tracts.
Lamina VI: Consists of pyramidal cells whose axons form
corticofugal fibers that project to subcortical structures (e.g.
corticothalamic projections).

90
PHYSIOLOGY
Dr. Basim Mohamad Awan Lecture 8
MOTOR FUNCTIONS OF THE CEREBRAL CORTEX
All voluntary movements involve the conscious activity of the "motor cerebral
cortex of the brain. The motor cortex lies in front of the central sulcus and
occupies most of the frontal lobe (fig. 13-1). It is divided into 4 separate areas:
[I] THE PRIMARY MOTOR AREA
[II] THE PREMOTOR AREA
[III] THE SUPPLEMENTARY MOTOR AREA
[IV] THE MOTOR ASSOCIATION AREA
[I] THE PRIMARY MOTOR AREA (Area 4)
The primary motor area of the cerebral cortex (area 4 in Brodmann classification)
occupies the precentral gyrus in the frontal lobe. The body is topographically
represented in an inverted and crossed manner (fig. 8-1). The upper part of the
face is bilaterally represented and the area of representation of each part is
proportionate to the degree of fine movements in this part, e.g. hands and muscles
of speech are represented by large areas, whilst the trunk is represented by a small
area. The primary motor area contains two types of neurons:
1. Dynamic neurons: Which discharge at high frequency for a short time at the
beginning of contraction causing the initial development of force?
2. Static neurons: Which discharge at a much slower frequency but for much
longer time to maintain contraction for as long as required.
FUNCTIONS OF THE PRIMARY MOTOR AREA
1. Initiation of voluntary, fine, discrete (separate) movements of the distal parts of
the body e.g. hands and fingers.
2. Facilitation of stretch reflex; i.e. facilitation of skeletal muscle tone and tendon
jerks.
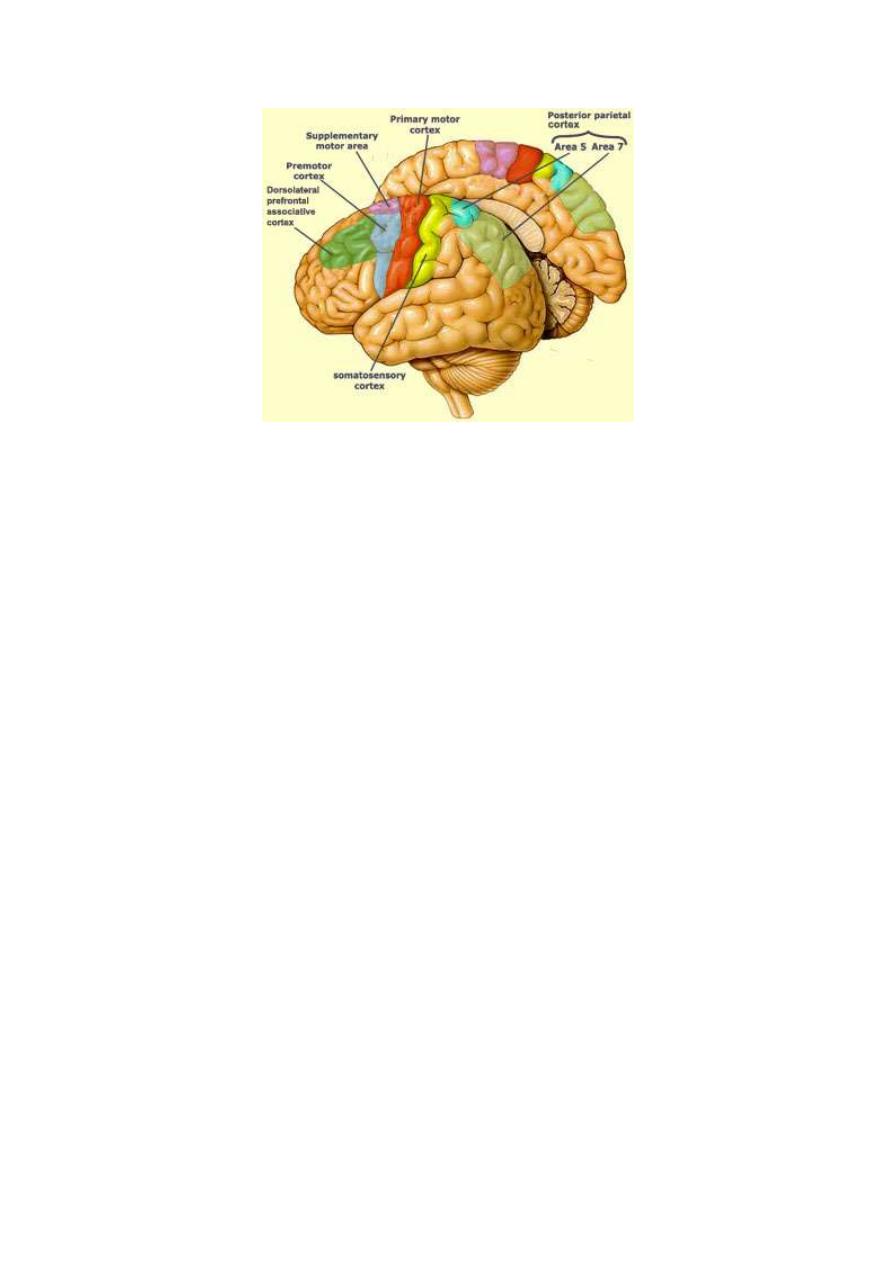
91
Figure 8 - 1: The lateral and medial surfaces of the left cerebral hemisphere showing the
primary motor cortex (M), the pre motor cortex (PM), the motor association area (MA)
and the supplementary motor area (SM).
[II] THE PREMOTOR AREA (Areas 6, 8 and 44)
The premotor area of the cerebral cortex (mainly area 6 of Brodmann
classification, but it also includes areas 8 and 44) lies immediately anterior to the
primary motor area. The topographic representation of the body is nearly the
same as in area 4. The premotor area includes some specialized areas with
specific functions (fig. 8-1):
1. Broca's area of speech (Word 44):
This area lies at the upper border of the lateral sulcus in front of the primary
motor cortex. It stores the motor programs for verbalization. Its damage leads to
inability to speak whole words except simple ones as "yes" or "no".
2. Eye field area (area 8):
It lies above Broca's area. It directs the eyes voluntarily towards any desired
object. It also controls the blinking movements of the eye lids. Damage of this
area leads to locking (fixation) of the eye on objects.
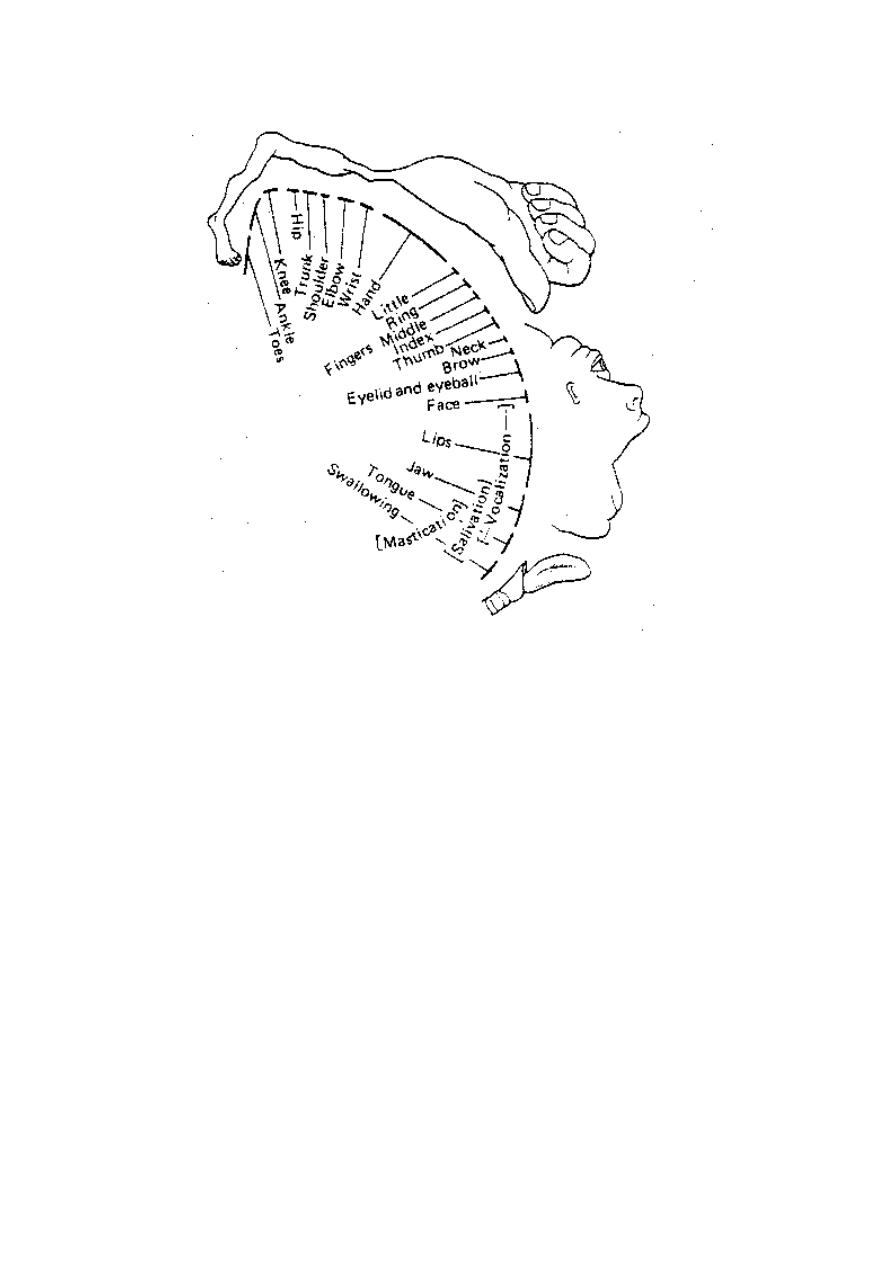
92
Figure 8-2: Body representation in the primary motor cortex
3. Head rotation area (part of area 6):
Lies immediately above area 8 and works in close association with it. It directs
the head towards different objects.
4. Hand skills area (part of area 6):
Lies immediately anterior to the primary motor area for hands and fingers. It
stores the motor programs for skilled hand movements; e.g. sharpening a pencil
or peeling a potato. Damage of this area leads to "motor apraxia"; i.e. inability
to do skilled hand movements. This area includes the "Exner center" for writing
skill.

93
FUNCTIONS OF THE PREMOTOR AREA
1. Initiation of gross movements that involve groups of muscles to support and'
facilitate fine movement, e.g. fix the shoulders and arms at a certain position so
that the hands and fingers can do skilled movements (e.g. threading a needle).
2. Weak inhibition of the stretch reflex. It tends to decrease the skeletal muscle
tone.
3. A center for rotation of the head towards objects.
4. A center for skilled hand movements.
5. A center for verbalization of words (area 44).
6. A center for voluntary eye and lid movements (area 8),
7. Inhibition of the grasp reflex.
THE MECHANISM OF ACTION OF THE PREMOTOR AREA
The premotor area acts by activating the corresponding nearby motor calls in me
primary motor area. This occurs in two ways:
First directly through direct projection fibers to area 4.
Second indirectly circuits starting from the premotor area to the corpus striatum
of the basal ganglia, to the thalamus, then back to terminate in area 4 of the
cerebral cortex.
[III] THE SUPPLEMENTARY MOTOR AREA
The supplementary motor area (fig. 8-1) is an extension of area 6 in the medial
surface of the cerebral hemisphere. Accordingly, it is also called "the medial
area 6". Topographic representation of the body is bilateral and in a horizontal
position; head anteriorly and legs posteriorly. This area is connected by
projection fibers to the premotor and the primary motor areas.
The supplementary motor area supplements the functions of the premotor area in
producing positioning and fixation of the different parts of the body as a

94
background for finer hand or feet movements; e.g. the coordinated movement of
the trunk with the hand and feet during boxing.
This area is involved in preparation for movements before they start. It s hows
electrical potentials shortly before the start of the movement (readiness
potential) and increase in metabolism and local blood flow on the mere intention
to move a muscle,
[IV] THE MOTOR ASSOCIATION AREA
This area occupies the frontal lobe in front of the premotor area (fig. 8-1). It
receives input signals from the parieto-tempqro-occipital association area and
projects output' signals to the motor and premotor areas through the caudate and
putamen circuits.
FUNCTIONS OF THE MOTOR ASSOCIATION AREA
1. Setting off goals and aims of movement and then taking the decision to
start the movement. Once the decision is taken, signals are sent to the basal
ganglia (the motor consultant of the cerebral cortex) to activate programs or set
plans for the movement. The motor plans and programs are then fed to the motor
and premotor areas.
2. Elaboration of thoughts; i.e. carrying out prolonged thought processes
which involve setting of plans and developing new constructive ideas. This is the
area of the brain which is concerned with deep quiet thinking during rest. A
lesion in this area abolishes creativity and planning for the future.
CONNECTIONS OF THE MOTOR CORTEX
AFFERENT CONNECTIONS
A. FROM OTHER CORTICAL AREAS
• Of the same side: from somatic sensory areas, visual and auditory areas.
• Of the opposite side: from the contralateral motor cortex to connect

95
corresponding points on both sides.
B. FROM THE THALAMUS
• The ventrobasal complex (VPL and VPM nuclei); it receives specific sensory
signals.
• The intralaminar nuclei; it receives nonspecific signals to arouse the cortex.
• The ventral and medial nuclei; it receives impulses coming from the
cerebellum and basal ganglia.
EFFERENT CONNECTIONS
A. Pyramidal tract fibers to motor nuclei in the brainstem and spinal
cord.
B. To the basal ganglia (the caudate and putamen circuits).
C. To the red nucleus of the midbrain.
D. To the cerebellum through the middle cerebellar peduncle
(cortico-ponto-cerebellar fibers).
E. To adjacent cortical areas to inhibit any unwanted discharge (lateral
inhibition). These inhibitory fibers are collaterals from the axons of the gi
ant
Betz cells. This helps to sharpen the outgoing signals.

96
THE MOTER TRACTS
Dr. Basim Mohamad
Alwan
Lecture 9
The motor tracts (also called the descending tracts) are divided into two groups of
tracts; the pyramidal and the extrapyramidal.
THE PYRAMIDAL TRACTS
The pyramidal tracts are three tracts of common origin but separate destinations
(fig. 9-1). These tracts are:
1. The corticonuclear tract; which originates in the cerebral cortex and terminates
on motor nuclei in the midbrain and pons.
2. The corticobulbar tract; which originates in the cerebral cortex and terminates
on motor nuclei in the pons and medulla.
3. The corticospinal tract; which originates in the cerebral cortex and terminates
on the motor neurons of the spinal cord.
THE ORIGIN OF THE PYRAMIDAL TRACTS
The pyramidal tracts originate from:
1. The primary motor area: 30% of the fibers.
2. The premotor and supplementary motor areas: 30% of the fibers.
3. The somatic sensory areas: 40% of the fibers.
The pyramidal tracts arise from lamina V of the cerebral cortex which contains
the pyramidal cells. There are two types of fibers in the pyramidal tracts:
a. Thick fibers (16 um in diameter). These arise from the giant pyramidal cells
(Betz cells) found only in the primary motor area. They constitute only 3% of the
total number of fibers.
b. Thin fibers (less than 4 um in diameter). These arise from small pyramidal
cells found in different parts of the cortex. They constitute the remaining 97% of
the fibers.

97
THE CORTICONUCLEAR TRACT
ORIGIN: From the eye field area in the frontal lobe (area 8) and the related areas in
the motor and the somatosensory areas.
COURSE AND DESTINATION:
Fibers descend down through the genu of the
internal capsule to the brainstem. They terminate on the nuclei of cranial nerves III,
IV in the midbrain and VI in the pons on both sides.
FUNCTIONS
1. Voluntary conjugate movements of the eye to look at different objects.
2. Facilitate the stretch reflex of the external ocular muscles.
THE CORT1COBULBAR TRACT
ORIGIN
:
From the lower part of the motor and sensory" areas of the cerebral cortex.
COURSE AND DESTINATION:
Fibers descend down through the genu of the
internal capsule to the pons and medulla oblongata. They cross to the opposite side
to terminate on the nuclei of the cranial nerves V, VII, IX, XI and XII.
FUNCTIONS
1. Voluntary movement of muscles in the head and neck.
2. Facilitation of stretch reflex of these muscles
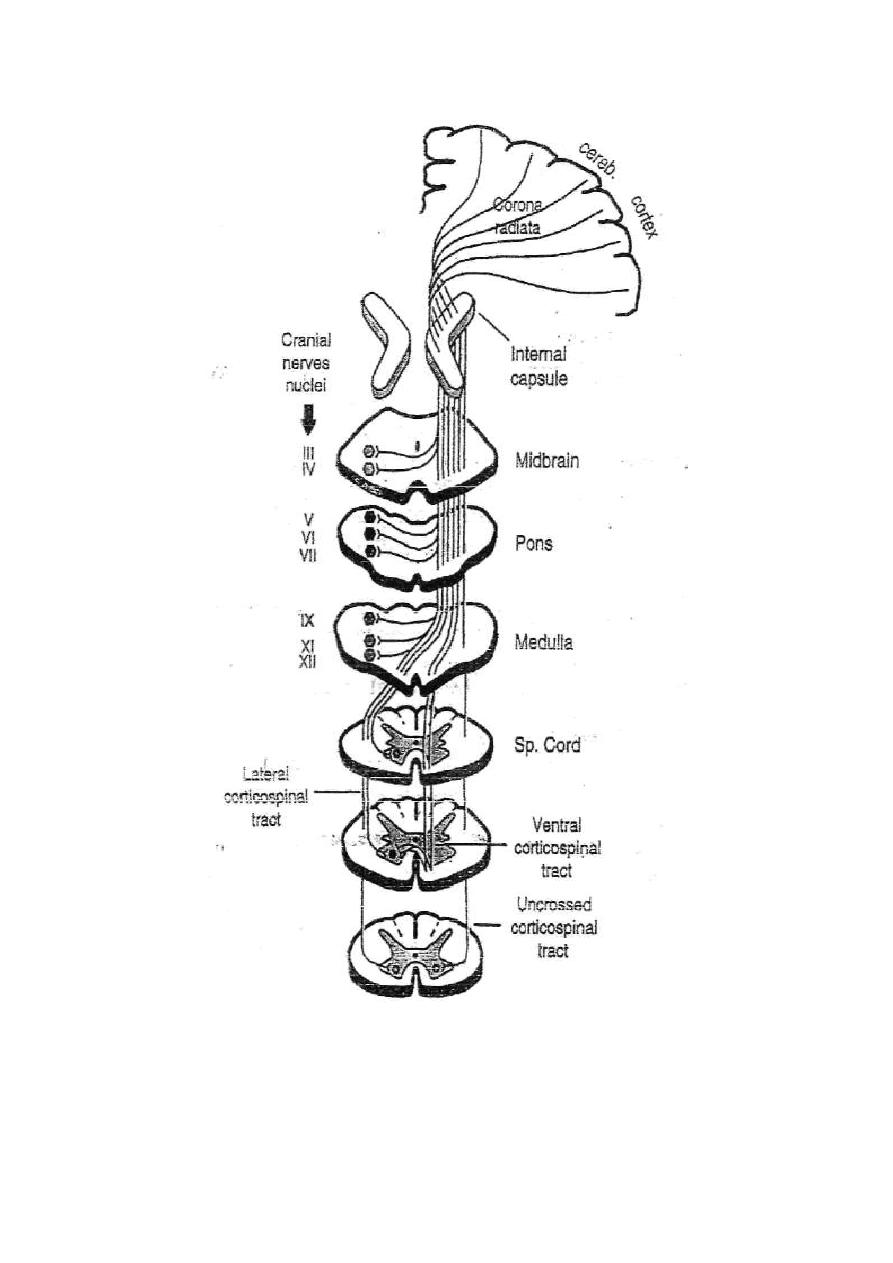
98
Figure 9 -1: The origin, course, and destinations of the pyramidal tracts.

99
THE CORTICOSPINAL TRACT
ORIGIN:
From the motor, premotor, supplementary motor and somatic sensory
areas of the cerebral cortex.
COURSE AND DESTINATIONS: Fibers descend in the corona radiata, then
through the anterior two-thirds of the posterior limb of the internal capsule down
to the brainstem. In the medulla, fibers collect together to form the medullary
"pyramid". In the lower medulla, pyramidal fibers take one of three courses:
1. 90% of the fibers cross to the opposite side in the motor decussation and
descend in the posterolateral column of the spinal white matter as the "lateral
corticospinal tract". They terminate on the ventral horn cells mainly through
intemeurons, but some fibers terminate directly on ventral horn cells.
2. 8% of the fibers descend directly in the ventral column of the spinal white
matter of the same side as the "ventral corticospinal tract". They cross
gradually as they descend in the cervical and upper thoracic segments of the
spinal cord to terminate on the ventral horn cells of the opposite side.
3. 2% of the fibers descend directly in the posterolateral column of the spinal
white matter as the "uncrossed corticospinal tract". They terminate on the
ventral horn cells of the same side.
FUNCTIONS
The crossed (lateral and ventral) corticospinal tracts have the following
functions:
1. Production of fine voluntary movements of the distal parts of the body e.g.
fingers and hands.
2. Facilitation of lower motor neurons and stretch reflex.

100
The uncrossed corticospinal tract has the following functions:
1. Provide bilateral innervation of some muscles as the respiratory and abdominal
muscles.
2. Gross positioning movements controlled by the supplementary motor area.
3. Help partial recovery of movements after injury of the crossed
corticospinal tracts.
THE EXTRA PYRAMIDAL SYSTEM
The extrapyramidal system (fig. 9-2) includes all parts of the nervous system other
than the pyramidal tracts that contribute to the control of skeletal muscle activity.
These parts include the basal ganglia, the red nucleus, the reticular formation of the
brainstem, the vestibular nuclei, and the tectum of the midbrain and the inferior olive
of the medulla. All these parts send signals down to the spinal motor nuclei in the
descending extrapyramidal tracts.
THE BASAL GANGLIA
It receives projection fibers from the motor cortex to corpus striatum and then to globus
pallidus then to several nuclei in the brainstem. These nuclei include the subthalamus,
substantia nigra, red nucleus, tectum of the midbrain, reticular formation, vestibular
nucleus and inferior olive.
THE RED NUCLEUS
It is located in the midbrain and receives projection fibers from the motor cortex (the
corticorubral tract) and collaterals from the corticospinal tract as it passes through
the midbrain. It also receives projection fibers from the globus pallidus of the basal
ganglia. All these fibers synapse in the lower part of the red nucleus which
contains giant pyramidal neurons similar to Betz cells. Rubrospinal tract
originates from these neurons, cross to the opposite side and descend in the spinal

101
cord very closely anterior to the lateral corticospinal tract. Fibers terminate on the
ventral horn cells either directly or through interneurons.
FUNCTIONS OF THE RED NUCLEUS
1. It is a relay station in the corticorubrospinal pathway which acts as an accessory
pathway for the corticospinal tracts. This pathway can initiate gross movements.
2. It is inhibitory to the motor neurons and stretch reflex through stimulation of
the inhibitory reticular formation of the brainstem.
Lesions of the pyramidal tracts usually involve the corticorubrospinal pathway. If the
latter is spared, a good deal of gross movements is retained. Only the fine
movements of the fingers and hands are considerably impaired.
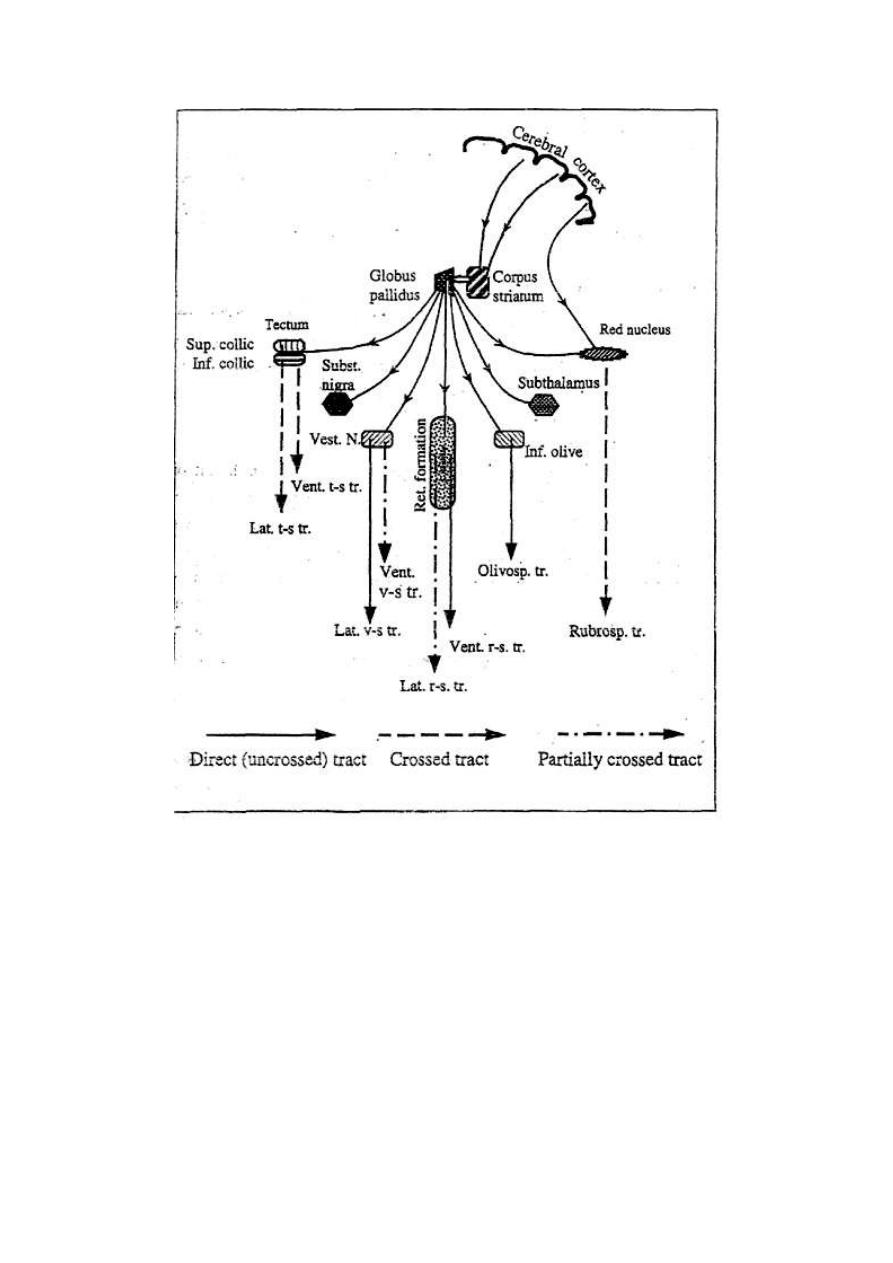
102
Figure 9 - 2: A schematic diagram of the extrapyramidal system.

103
THE TECTUM OF THE MIDBRAIN
It receives projection fibers from the globus pallidus of the basal ganglia, and
gives origin to two descending extrapyramidal tracts:
* The lateral tectospinal tract: Originates from the superior colliculus (the
center of visual reflexes), crosses to the opposite side and terminates in the cervical
segments of the spinal cord. It is concerned with directing the eye and turning the
head towards a light source (visuospinal reflexes).
* The ventral tectospinal tract: Originates from the inferior colliculus (the
center of auditory reflexes), crosses to the opposite side and terminates in the
cervical segments of the spinal cord. It is concerned with turning the head to direct
the ears towards a sound source (audiospinal reflexes).
THE RETICULAR FORMATION OF THE BRAINSTEM
It receives projection fibers from the globus pallidus of the basal ganglia, and
gives origin to two descending extrapyramidal tracts:
* The lateral reticulospinal tract: Originates from the inhibitory reticular
formation of the medulla. Some fibers cross to the opposite side, but most fibers
descend in the same side of the spinal cord. It inhibits the gamma motor neurons,
thus inhibiting the stretch reflex and skeletal muscle tone.
• The-ventral reticulospinal tract: Originates from the facilitatory reticular
formation of the pons. Fibers descend without crossing to terminate on the gamma
motor neurons of the ipsilateral side of the spinal cord. It facilitates the gamma motor
neurons, thus facilitating the stretch reflex and the skeletal muscle tone.
THE VESTIBULAR NUCLEUS OF THE MEDULLA
It receives projection fibers from the globus pallidus of the basal ganglia, and
gives origin to two descending extrapyramidal tracts;
• The lateral vestibulospinal tract: Originates from the vestibular

104
'nucleus, descends uncrossed to terminate on the alpha and gamma motor neurons.
• The ventral vestibulospinal tract: Originates from the vestibular
nucleus, descends on both sides of the spinal cord to terminate on
the alpha and gamma motor neurons.
The vestibulospinal tracts facilitate the stretch reflex and skeletal muscle tone.
They mediate some postural reflexes
THE INFERIOR OLIVE OF THE MEDULLA
The inferior olive receives input fibers from the motor cortex, the globus pallidus
of the basal ganglia and the spinal cord. It sends output fibers to the cerebellum. It
projects the fibers of the olivospinal tract which descend in the spinal cord to
terminate on the ventral horn cells of the same side. The olivospinal tract is
facilitatory to the stretch reflex and the skeletal muscle tone.
The inferior olive works in close association with the cerebellum to correct any
deviation of the muscle contraction from the preset plan of movement (the
servocomparator function of the cerebellum).
GENERAL FUNCTIONS OF THE EXTRAPYRAMIDAL SYSTEM
1. Mediation of gross movements which involve a group of large muscles.
2. Provides a weaker alternative to the pyramidal system for mediation of some
discrete movements.
3. Mediation of fixation and positioning movements which accompany other fine
movements.
4. Adjustment of the skeletal muscle tone through facilitation or inhibition.
5. Adjustment of muscle movements to match preset plans to reach a certain target.

105
THE UPPER AND LOWER MOTOR NEURONS
To do a voluntary movement, signals start in the motor neurons of the cerebral
cortex and reach the skeletal muscles through two orders of neurons:
1. Upper motor neurons:
These are the neurons of the pyramidal and extrapyramidal tracts in the CNS. They
extend from the cerebral cortex and the extrapyramidal nuclei down to the motor
neuron pool of the brainstem and spinal cord. Neurons of the motor neuron pool
themselves are not included in the upper motor neurons.
2. Lower motor neurons:
These are the neurons of the motor neuron pool and their axons which form the
motor nerves to the skeletal muscles.
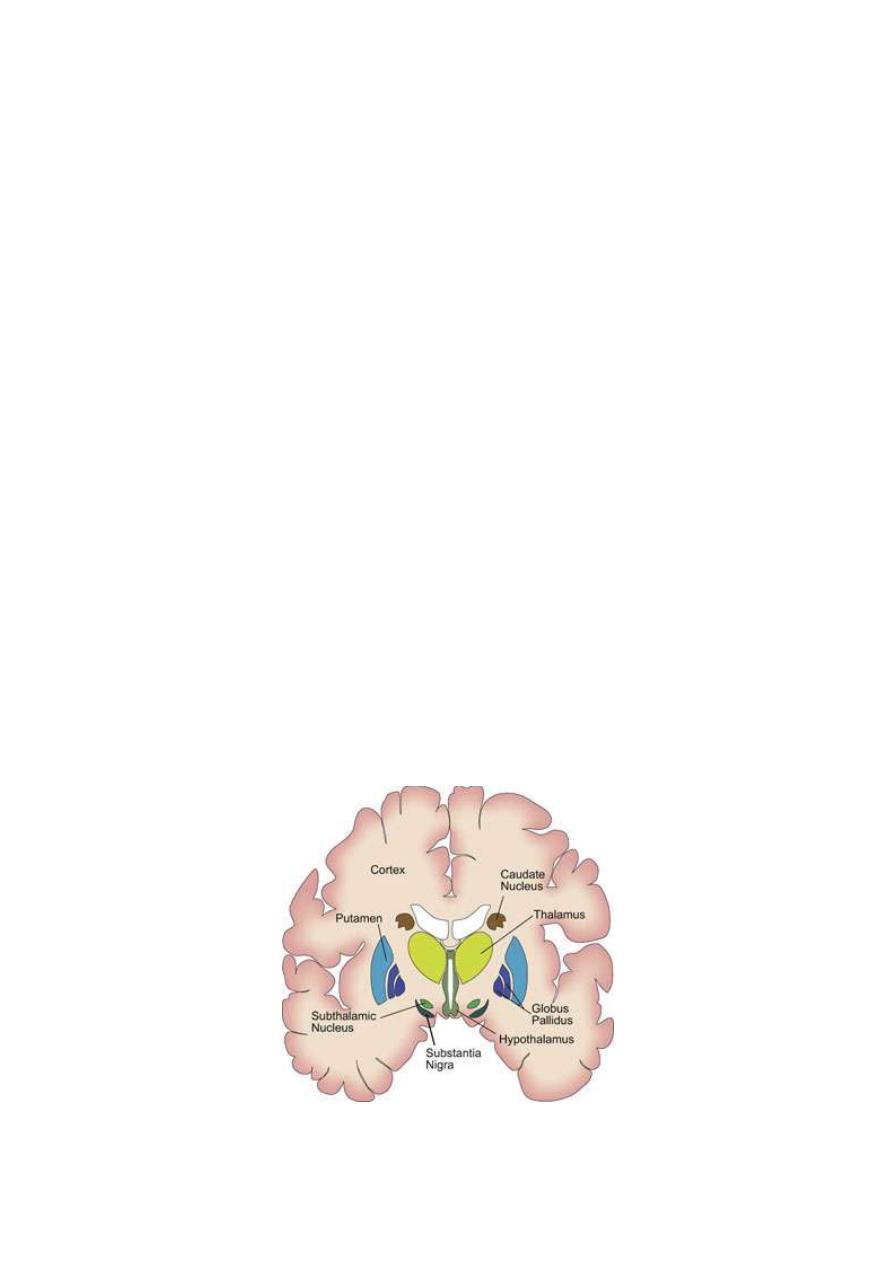
106
THE BASAL GANGLIA
Dr. Basim Mohamad Alwan Lecture 10
The basal ganglia (fig. 10-1) are sub cortical masses of gray matter found in the cerebral
hemispheres lateral to the thalamus that includes:
1. The caudate nucleus.
2. The lentiform (lenticular) nucleus. This consists of 2 parts:
a. An outer part called the putamen.
b. An inner part called the globus pallidus , which further divided into external
and internal segments.
Both caudate nucleus and putamen are called the corpus striatum (striate body or
striatum).
3. The subthalamic nucleus (subthalamus or body of LUYS).
4. The substantia nigra in the midbrain.
The afferent connections to the basal ganglia terminate mainly in the corpus
striatum, whilst the efferent connections originate mainly from the globus pallidas.
Figure 10-1 Transverse section in the cerebrum showing the basal ganglia
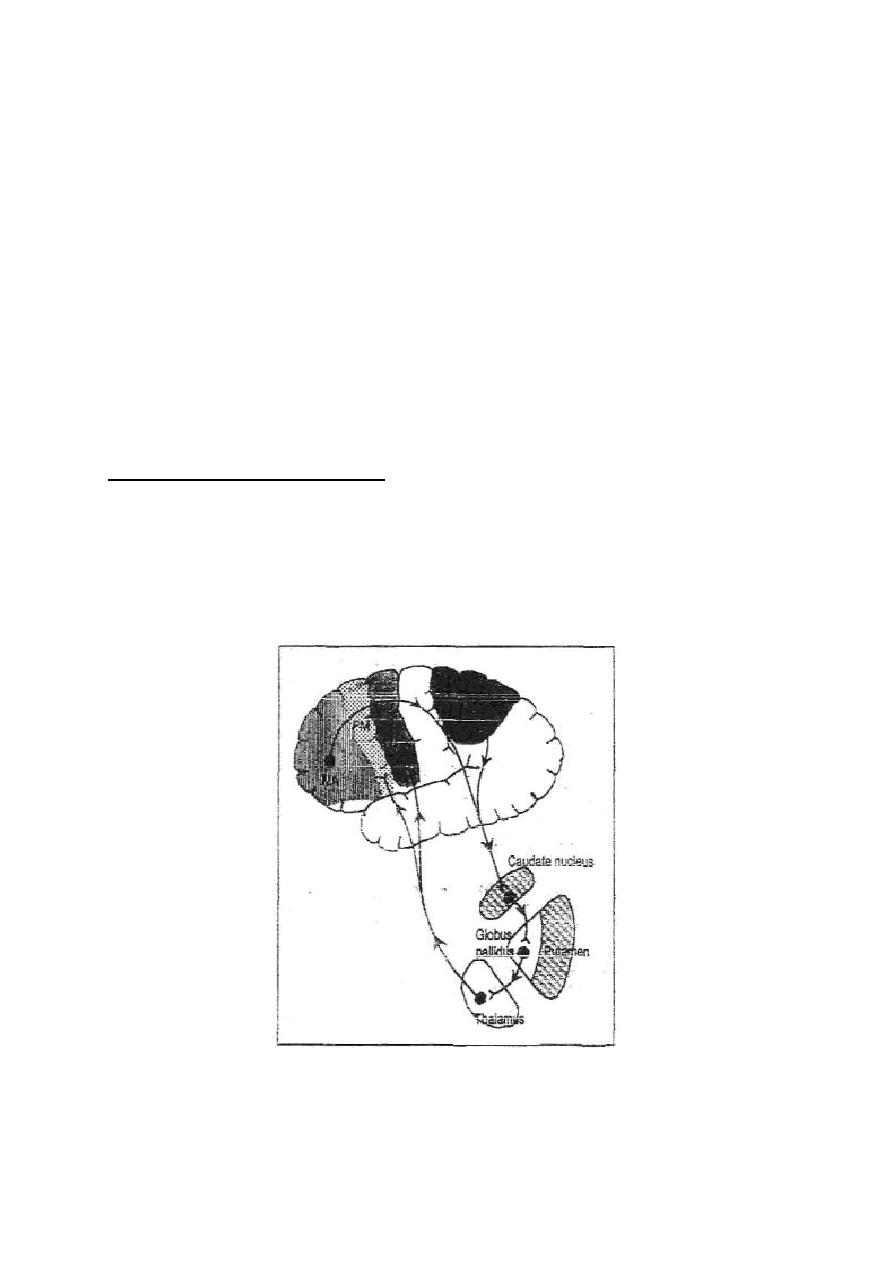
107
NEURAL CONNECTIONS OF THE BASAL GANGLIA
There are numerous free interconnections between various nuclei of the basal
ganglia. This is in addition to the connections with the cerebral cortex, thalamus
and brainstem.
CONNECTIONS WITH THE CEREBRAL CORTEX
The basal ganglia are connected to the cerebral cortex through two main circuits,
the caudate and putamen circuits.
THE CAUDATE CIRCUIT
The caudate circuit (fig.10-2) starts at the motor association and sensory
association areas of the cerebral cortex → to the caudate nucleus → the globus
pallidas → ventrolateral (VL) nucleus of the thalamus → premotor and primary
motor areas of the cortex.
Figure 10-2: The caudate circuit
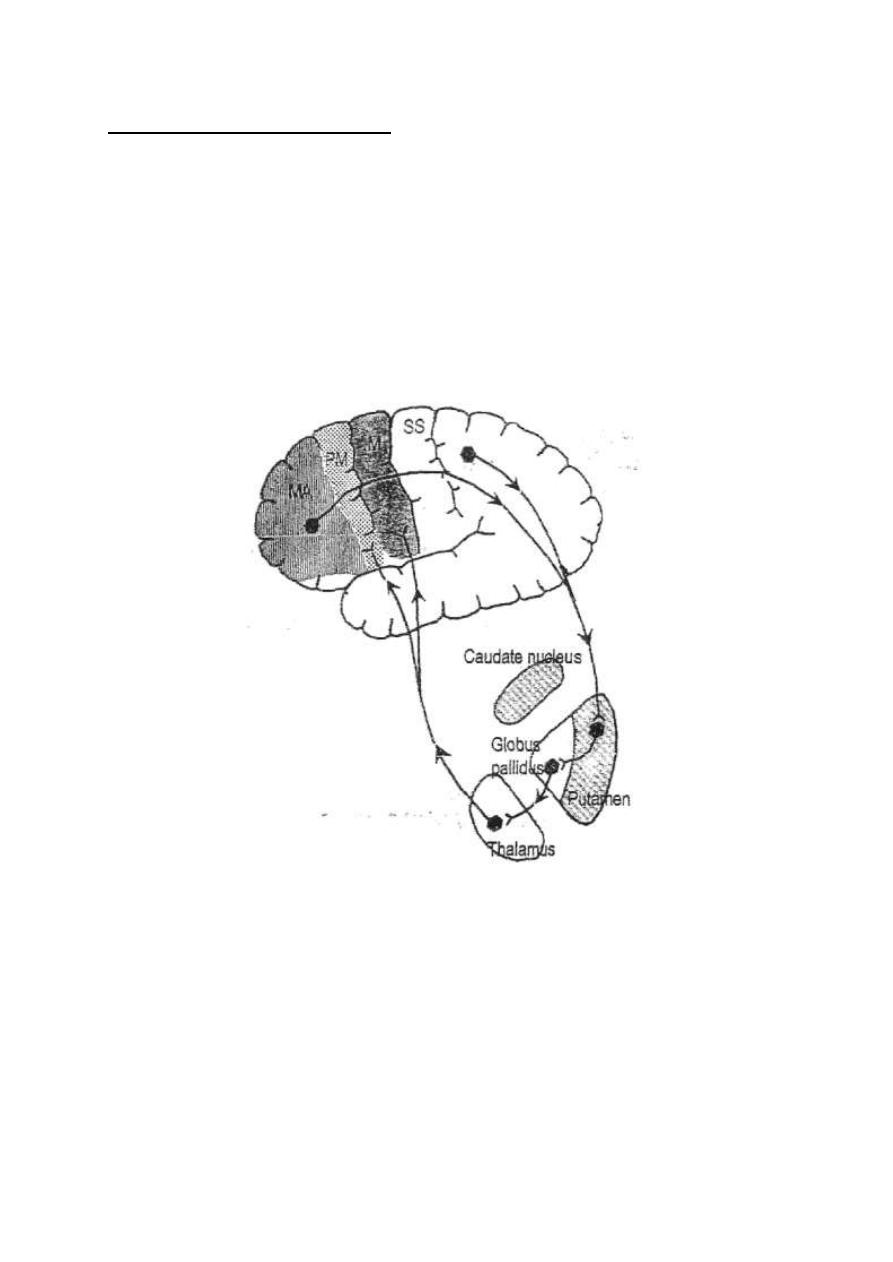
108
THE PUTAMEN CIRCUlT
The putamen circuit (fig.10-3) starts at the motor association and sensory
association areas → to the putamen → the globus pallidus → the ventrolateral
(VL) nucleus of the thalamus → premotor and primary motor areas of the cortex.
There are other circuits which are closely associated with the putamen circuit.
They connect the basal to the subthalamus, substantia nigra:
(a) Putamen → globus pallidus → subthalamus → thalamus → motor cortex.
(b) Putamen → globus pallidus → substantia nigra→thalamus→ motor cortex
Figure 10- 3: The putamen circuit.
Through these circuits the basal ganglia receive thoughts and ideas from the
cerebral cortex and feedback plans and programs for motor actions to the motor
and premotor areas.
CONNECTIONS WITH THE BRAINSTEM
The basal ganglia are connected with some centers in the brainstem, from which

109
fibers descend in the extrapyramidal tracts down to the spinal cord. Fibers project
from the globus pallidas to:
1. The reticular formation, from which the reticulospinal tracts originate.
2. The red nucleus, from which the rubrospinal tract originates.
3. The vestibular nucleus, from which the vestibulospinal tracts originate.
4. The inferior olive, from which the olivospinal tract originates.
This is in addition to fibers to the subthalamic, substantia nigra, and tectum.
NEUROTRANSMITTERS IN THE BASAL GANGLIA
• Fibers from the cerebral cortex to the corpus striatum release
glutamate, Intrastriatal intemeurons release acetylcholine when
stimulated by glutamate.
• Fibers from the substantia nigra to the corpus striatum release dopamine.
• Fibers from the corpus striatum to the globus pallidus and substantia nigra
release gamma-aminobutyric acid (GABA).
• Fibers from the brainstem to the basal ganglia release noradrenaline,
serotonin and enkephalin.
Glutamate, acetylcholine and noradrenaline are excitatory transmitters to the
basal ganglia, whilst dopamine, GABA, serotonin and enkephalin are inhibitory.
A delicate balance between the excitatory and inhibitory transmitters is essential
for proper functioning of the basal ganglia.
METABOLIC CHARACTERISTICS OF THE BASAL GANGLIA
The basal ganglia are characterized metabolically by the following:
1. High metabolic rate, with high rate of O
2
consumption. They suffer rapidly
from hypoxia or ischemia.
2. High ability to concentrate copper. Excess copper in the plasma is bound to a

110
protein made in the liver called ceruloplasmin until it is excreted in bile.
In Wilson disease, there is failure, of ceraloplasmin formation and inability to
excrete copper in bile. Serum copper level rises which leads to copper
intoxication. Large amounts of copper accumulate in the cells of the liver and the
lentiform nucleus of the basal ganglia which lead finally to hepatolenticular
degeneration.
FUNCTIONS OF THE BASAL GANGLIA
The functions of basal ganglia are purely motor. In lower animals and birds in
which the cerebral cortex is poorly developed, the basal ganglia act as the primary
motor cortex. In humans, they play an important role in controlling the muscle
tone and voluntary movements'.
[A] ROLE IN CONTROLLING THE MUSCLE TONE
The caudate nucleus stimulates muscle tone through stimulation of the vestibular
nucleus and inferior olive.
The lentiform nucleus inhibits the muscle tone through inhibition of the primary
motor cortex and stimulation of the inhibitory reticular formation and red nucleus
of the brainstem.
The inhibitory influence of the basal ganglia on the muscle tone is stronger than
the stimulatory influence.
[B] ROLE IN CONTROLLING VOLUNTARY MOVEMENT
Planning and programming of movements start in the basal ganglia. Neurons of
the basal ganglia discharge before the movement begins. The basal ganglia works
in collaboration with the cerebral cortex through the caudate and putamen circuits
to produce a coordinated, organized and purposeful movement.

111
ROLE OF THE CAUDATE CIRCUIT
1. Design of plans which convert thoughts and ideas into motor actions: The
caudate circuit directs the motor activity according to the thoughts of mind; i.e.
converts thoughts into motor plans. It determines what patterns of movements
will be used and in what sequence to achieve a complex goal; e.g. when dressing,
one puts on the shirt then the necktie before the jacket.
2. Determining the timing and scale of movement; The basal ganglia
determine:
(a) To what extent the movement will be fast.
(b) For how long the movement will last.
Damage of the caudate circuit leads to:
• Inability to organize movements to achieve a complex goal; e.g. one wears the
necktie before the shirt.
• Inability to write or draw figures with a fixed scale; e.g. if a patient
with a lesion in the left caudate circuit draws a face of a human
being, he draws the left side of the face (which is in the right field of
vision) to a much smaller scale than the right side (fig. 10-4).
• Excluding the contralateral side of the body from any motor plans.
The patient avoids using the contralateral side of his body and ignores it as if it
does not exist.
E.g. when the patient walks he swings the arm of the healthy side
but not that of the ignored side.

112
Figure 10 - 4: A typical drawing made' by patient with a lesion in the left
basal ganglia or the left parietal lobe of the cerebral cortex.
ROLE OF THE FUTAMEN CIRCUIT
Storage of motor programs of familiar motor actions: e.g. writing
one's name or signature, lighting a candle or a kerosene lamp.
Damage of this circuit leads to motor apraxia ; i.e. loss of ability to
carry out familiar movements in the absence, of muscle paralysis. E.g.,
one finds difficulty to write as if he is learning to write for the first time.
MANIFESTATIONS OF LESIONS IN THE BASAL GANGLIA
Lesions in the basal ganglia produce two characteristic features which appear on
the contralateral side of the lesion:
1. Involuntary movements during rest. These movements are in the form of
chorea, athetosis, hcmiballismus or static tremor. The movements disappear during

113
sleep but increase with nervous excitement.
2. Change in muscle tone; either hyper or hypotonia.
(A) CHOREA (Gr.: choreia means dance)
CAUSE: Lesion in the caudate nucleus.
FEATURES:
• Involuntary, rapid, purposeless "dancing" movements during rest. They might
be superimposed on voluntary movements leading to their disruption.
• Hypotonia (caudate nucleus is normally facilitatory to the stretch reflex).
• Chorea is accompanied by loss of the caudate circuit functions (see above).
TYPES OF CHOREA
1. Sydenham chorea: Which occurs in young children (5-15 years) as a
complication of rheumatic fever? It is more common in females than in males.
2. Huntington chorea: Which is a hereditary disease that appears usually at the
fourth decade of life? It is accompanied with, dementia and block of much of the
thinking processes.
3, Chorea gravidarum: which occurs during pregnancy and chorea of the
contraceptive pills?
(B) ATHETOSIS (Gr.: athetos means not fixed)
CAUSE: Lesion in the putamen.
FEATURES:
Involuntary, spasmodic, slow, writhing (snake-like) movements of one or
more limbs, particularly the hands. It is also called "mobile spasm"
because the limbs cannot be kept at a fixed position.
• Hypertonia (putamen is normally inhibitory to the stretch reflex).
• Athetosis is associated with loss of putamen circuit functions.

114
(C) HEMIBALL1SMUS (Gr.: ballismus means jumping)
CAUSE: Lesion in the subthalamus.
FEATURES:
•
Sudden, involuntary, strong, spasmodic movements that involve
one limb or a whole one side of the body. It might occur in one
leg during walking leading to stumbling of the body.
(D) PARKINSONISM (Parkinson disease, or Paralysis agitans)
CAUSE: Lesion in the substantia nigra leading to degeneration of the
dopaminergic neurons that project to the corpus striatum.
Normally, there is a steady loss of dopaminergic neurons and receptors with age.
This process is accelerated by atherosclerosis or phenothiazine tranquilizers
which block dopamine receptors. In this case, an imbalance occurs between
dopamine (from the nigral projection fibers) and acetylcholine (from the cortical
projection fibers) in the corpus striatum.
FEATURES:
Three features characterize parkinsonism; tremor, rigidity and akinesia.
1. TREMOR
Tremor is an involuntary, rhythmic, oscillating movement due to rhythmic short
contractions of muscles. The tremor of Parkinsonism occurs at a frequency of 4-8
Hz. At the hands, it usually takes the form of pill-rolling movement. The tremor
might occur in the muscles of mastication leading to an up and down movement
of the mandible (mandibular tremor). The tremor occurs in the waking time
during rest. It markedly diminishes or disappears during voluntary movement.
That is why it is called a "static tremor". It disappears also during sleep.
The parkinsonian tremor is caused by activation of negative feedback circuits
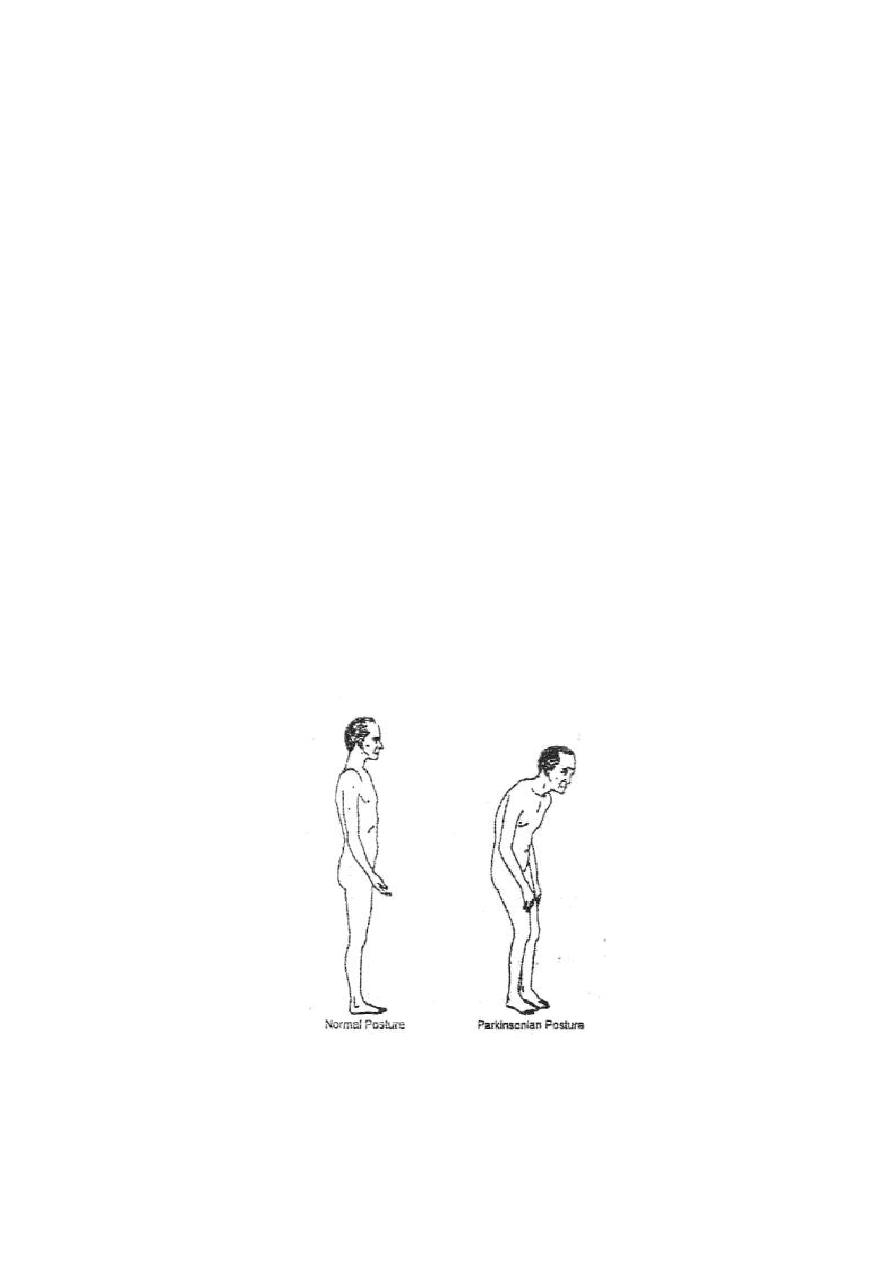
115
between the basal ganglia and the motor cortex → rapid on-off inhibition of the
motor cortex → alternating activation inactivation of the muscles → tremor.
2. RIGIDITY
Parkinsonian rigidity occurs in both flexor and extensor muscles but is more in the
flexors leading to a flexor attitude of the patient (fig. 10-5). When bending a limb
of a parkinsonian patient, the muscle rigidity resists the bending all the way
during bending, making the limb appearing as a pipe of lead (lead pipe rigidity),
or it may give a series of "catches" or "clicks' during the bending (cog-wheel
rigidity).
Parkinsonian rigidity is caused by:
(a) Facilitation of the gamma motor neurons due to lo
SS
of the supraspinal
inhibitory effect of the lentiform nucleus.
(b) Facilitation of the alpha motor neurons due to lack of dopamine. The caudate
nucleus becomes hyperactive. It sends facilitatory discharge via the
extrapyramidal tracts down to the alpha motor neurons of the motor neuron pool.
Figure 10-5: The change in posture in case of Parkinsonism.

116
3. AKINESIA
Akinesia means lack of movement. Each movement is carried out as a "sequence"
of phases of excitation and inhibition. Due to lack of dopamine, the basal ganglia
is "locked" at a state of excitation. So, it cannot make a plan or activate a program
of a movement. The patient avoids voluntary movements because he finds great
difficulty in initiating this movement due to the absence of the planning and
programming function of the basal ganglia.
Parkinsonian akinesia is manifested by:
(a) Lack of voluntary movement due to difficulty to initiate this movement.
The patient appears as if he is paralyzed although he is not. That is why
Parkinson disease is also called "paralysis agitans".
(b) Refraining from habitual movements, e.g. rolling of beads, or
manipulating one's hairs during reading.
(c) Absence of associating habitual movements, e.g. swinging of arms during
walking.
(d) Mask face; due to lack of movement of facial muscles of expression,
(e) Slow, monotonous, low-volume speech.
(f) The gait (the way of walking) is shuffling; the patient walks in short steps
without lifting his feet off the ground.
TREATMENT OF PARKINSONISM
Medical: As Parkinsonism is caused by an imbalance between dopamine and
acetylcholine in the basal ganglia, it is treated by either reducing acetylcholine
activity or increasing dopaminergic activity. Reducing Ach activity is done by
giving atropine (a muscarinic cholinergic blocker). Increasing dopaminergic
activity is done by giving L-dopa. L-dopa is preferred over dopamine because
L-dopa crosses the blood-brain barrier much easier.

117
Surgical: Destruction of the ventrolateral nuclei of the thalamus by
electrocoagulation disrupts the feedback circuits between the basal ganglia and
the motor cortex. This abolishes the tremor.

118
Dr. Basim Mohamad Alwan Lecture 11
THE CEREBELLUM
The cerebellum receives direct or indirect input signals from all the
sensory pathways. This sensory information does not reach conscious
perception. The cerebellum functions as a special kind of computer
which automatically integrates the sensory inputs to provide rapid
responses of smooth coordinated motor actions.
FUNCTIONAL ANATOMY
Cerebellum (fig. 11-1, 11-2) consists of a midline structure called the vermis,
two cerebellar hemispheres on both sides, and a separate lobe called the
flocculonodular lobe on the inferior surface.
Three pairs of cerebellar peduncles connect the cerebellum with the brainstem.
The superior cerebellar peduncle (SCP) connects the cerebellum with "the
midbrain, the middle peduncle (MCP) with the pons and the inferior peduncle
(ICP) with the medulla.
PHYSIOLOGICAL DIVISIONS OF THE CEREBELLUM
Physiologically, the cerebellum is divided into three functional divisions (fig.
16-1):
1. THE ARCHICEREBELLUM (vestibulocerebellum); is the oldest part of the
cerebellum. It consists of the flocculonodular lobe. It is connected to the
vestibular apparatus through the vestibular nuclei in the medulla. It is concerned
with maintenance of equilibrium.

119
Figure 11 - 1: The anatomical (right) and functional (left) divisions of the
cerebellum.
2. THE PALEOCEREBELLUM (spinocerebellum); which consists of the vermis and
the medial paravermal parts of the cerebellar hemispheres. It receives signals from
the peripheral proprioceptors. It is concerned with the control of muscle tone and
gross movement.
3. THE NEOCEREBELLUM (cerebrocerebellum or the pontocerebelium); which
consists of the lateral parts of the cerebellar hemispheres. It is connected to the
cerebral cortex. It is concerned with the control of skilled voluntary movements
initiated by the cerebral cortex.
All these three divisions are interconnected by interneurons.

120
Fig. 11-2 Functional parts of the cerebellum
THE STRUCTURE OF THE CEREBELLUM
A section in the cerebellum shows an outer layer of gray matter which forms the
cerebellar cortex and an inner core of cerebellar white matter. The cerebellar cortex
is made up of three layers (fig. 11-3):
1. An outer molecular layer containing mainly interconnecting fibers.
2. An intermediate Purkinje cell layer containing mainly the giant Purkinje
cells.
3. An inner granule cell layer containing mainly granule cells.
The cortices of the various parts of the cerebellum are freely interconnected by
interneurons.
The deep white matter contains four cerebellar nuclei (fig. 11-6).
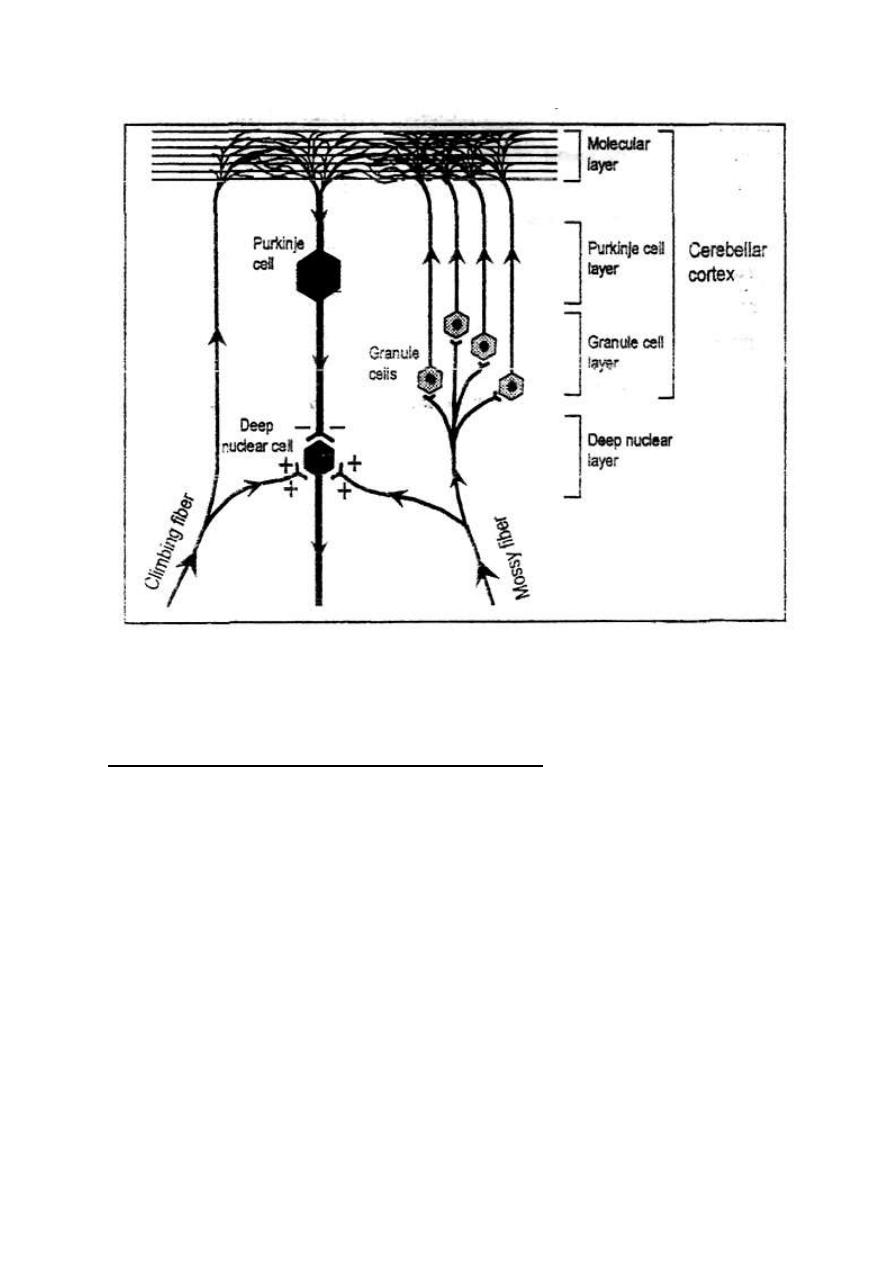
121
Figure 16-3: Cellular layers of the cerebellum. The climbing and Mossy
fibers
AFFERENT FIBERS TO THE CEREBELLUM
All the afferent fibers to the cerebellum are one of two types (fig. 11-3):
(A) CLIMBING FIBERS: Which originate from the inferior olive. They enter
the cerebellum, give off collaterals to the nuclear cells and proceed to terminate
on the dendritic branches of the Purkinje cells in the molecular layer of the cortex.
(B) MOSSY FIBERS: Which originate from all parts of the CNS including me
inferior olive. They give off collaterals to the nuclear cells and terminate on the
granule cells of the cerebellar cortex. The granule cells then relay on the dendritic
branches of the Purkinje cells in the molecular layer of the cortex.

122
All the afferent fibers to the cerebellum terminate in the cortex. All parts of the
cerebellar cortex send inhibitory projection fibers from the Purkinje cells to the
deep nuclear cells. These fibers are the only projection fibers from the cortex.
Some parts of the cortex of the flocculonodular lobe send direct projection fibers
to the vestibular nucleus in the medulla oblongata. In this respect, the vestibular
nucleus functions as a cerebellar nucleus.
The efferent fibers from the cerebellum are the axons of the nuclear cells. The
nuclear cells receive continuous excitatory input from the mossy fibers and
inhibitory input from the Purkinje cells. The net effect is moderate continuous
facilitation leading to continuous tonic facilitatory discharge from deep nuclear
cells during rest. Stimulation of nuclear cells
excitatory output signal (i.e.
increase in the frequency of discharge in the efferent fibers). Inhibition of nuclear
cells
inhibitory output signal
(i.e. decrease in the frequency of discharge in
the efferent fibers).
CEREBELLAR CONNECTIONS
AFFERENT CONNECTIONS (fig. 11 - 3)
[I] FROM THE BRAIN
1. TECTO-CEREBELLAR FIBERS: They originate from the superior and inferior
colliculi in the tectum of the midbrain
Sup. cer. ped.
different parts of the
ipsilateral cerebellum.
2. CORTICO PONTO CEREBELLAR FIBERS: They originate from the premotor area
of the cerebral cortex
pontine relay nuclei
Mid. cer. ped.
contralateral
cerebrocerebellum. They transmit signals from the cortex giving an "efference
copy" of the plan of the intended movement.

123
3. OLIVO CEREBELLAR FIBERS: They originate in the inferior olive
inf. cer.
ped.
all parts of the ipsilateral cerebellum. Inferior olive receives fibers from the
motor cortex, basal ganglia, reticular formation and spinal cord. The inferior olive
compares the signals from the brain with those from the spinal cord and feed the result to
the cerebellum.
4. VESTIRULO CEREBELLAR FIBERS: They originate from the vestibular nucleus
inf. cer. ped.
the ipsilateral vestibule cerebellum. They
transmit signals about body posture and equilibrium.
5. RETICULO CEREBELLAR FIBERS: They originate from the
reticular formation
mid. and inf. cer. ped.
the ipsilateral
spinocerebellum. They transmit signals of various sensations.
124
Figure 11-4: The afferent cerebellar connections.
[II]FROM THE PERIPHERY
The cerebellum receives input signals from all sensations. Most of the sensory
input comes through collaterals from the sensory pathways to the sensory
reticular formation and from there on to the cerebellum via the reticulocerebellar
fibers.
Proprioceptive informations are continuously carried to the cerebellum via the
dorsal and ventral spinocerebellar pathways.
1. THE DORSAL SPINOCEREBELLAR PATHWAY
This pathway (fig. 11-5) consists of two orders of neurons.
125
FIRST ORDER NEURONS: are the dorsal root neurons. Their afferent type-II
fibers enter the spinal cord in the dorsal root
terminate in the Clarke nucleus
(lamina VI of the spinal gray matter).
SECOND ORDER NEURONS:
are those of Clarke nucleus at the base of the
dorsal horn. Their fibers ascend in the same side up in the posterolateral column
of the spinal white matter as the dorsal spinocerebellar tract up to the medulla
oblongata. In the medulla fibers pass through the inferior cerebellar peduncle to
terminate in the cortex of the spinocerebellum.
Figure 11 - 5: The dorsal spinocerebellar pathway.
FUNCTIONS: It transmits static and dynamic proprioceptive signals from the
muscles, tendons and ligaments to the cerebellum.
126
2. THE VENTRAL SPINOCEREBELLAR PATHWAY
The ventral spinocerebellar pathway (fig. 11-5) consists of two orders of neurons.
FIRST ORDER NEURONS: are the dorsal root neurons. Their afferent type-II
fibers enter the spinal cord in the dorsal root and then terminate in lamina VII of
the spinal gray matter.
SECOND ORDER NEURONS: are neurons of lamina VII of the spinal gray
matter. Some of the fibers cross to the opposite side, the others ascend in the same
side up in the anterolateral column of the spinal white matter as the ventral
spinocerebellar tracts up to the midbrain. In the midbrain, fibers of the ventral
spinocerebellar tracts of both sides turn to pass through the superior cerebellar
peduncle to terminate in the cortex of the spinocerebellum.
Figure 11- 6: The ventral spinocerebellar pathway.

127
FUNCTIONS
:
Neurons of the ventral spinocerebellar tract are stimulated by
signals reaching the ventral horn cells of the spinal cord in the motor tracts. It
informs the cerebellum about the motor signals reaching the ventral horn cells.
So, it gives the cerebellum an "afference copy" of the final motor orders which
will be executed. The pathway also conducts some proprioceptive information
from the periphery to the cerebellum. But in that respect it is less important than
the dorsal pathway.
EFFERENT CONNECTIONS (fig. 11-6)
All the efferent (output) fibers from the cerebellum arise from the deep cerebellar
nuclei. They are axons of the deep nuclear cells.
[I] FROM THE ARCHICEREBELLUM
Fibers originate in the fastigial nucleus in the flocculonodular lobe
inf. cer.
ped.
vestibular nucleus. Some fibers arise from the cortex of the
flocculonodular lobe and project directly to the vestibular nucleus. Others
originate in the fastigial nucleus and descend to the spinal cord (fastigiospinal
tract).
[II] FROM THE PALEOCEREBELLUM
Fibers originate in two nuclei:
1. Fastigial nucleus in "the vermis.
inf. cer. ped.
reticular formation.
2. Nucleus interpositus in the paravermal portion:
•
Sup. Cer. Ped.
ventral thalamic nuclei -» motor cortex
•
Sup. Cer. Ped.
medial thalamic nuclei -» basal ganglia.
•
- Sup. Cer. Ped.
red nucleus.
•
Sup. Cer. ped.
culomotor nucleus
•
Inf. Cer. Ped.
vestibular nucleus.
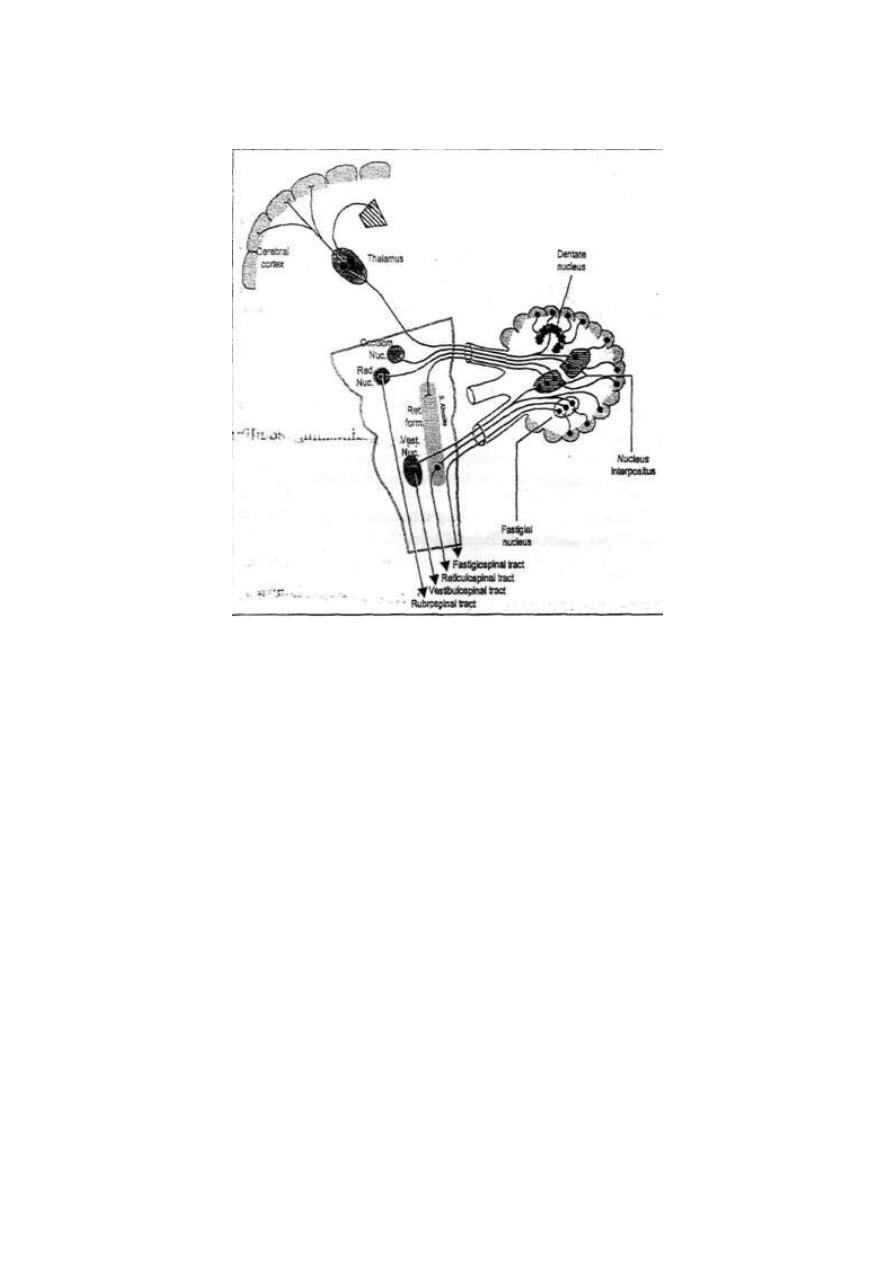
128
Figure 11 - 7: The efferent cerebellar connections
[II] FROM THE NEOCEREBELLUM
Fibers originate in the dentate nucleus in the cerebellar hemisphere
sup. cer. ped.
then cross to the opposite side in the brainstem
ventrolateral nucleus of the
thalamus
cerebral sensorimotor cortex.
The cerebellum is connected to the cerebral cortex by afferent and efferent fibers.
These form a neuronal circuit that start and end in the cerebral cortex. This circuit is
called the cortieo ponto cerebello dentate thalamo cortical circuit.

129
Physiology
FUNCTIONS OF THE CEREBELLUM
Dr. Basim Mohamad Alwan Lecture 12
[1] CONTROL OF POSTURE AND EQUILIBRIUM
This is mainly the function of the archicerebellum and some portions of the
vermis. The cerebellum plays a minor role in maintaining posture through a
servocomparator function. It compares signals from the vestibular apparatus (i.e.
the nonauditory labyrinth) with proprioceptive signals from different parts of the
body then sends corrective signals to adjust muscle tone to maintain posture. The
same mechanism occurs to maintain equilibrium during slow movement.
The cerebellum plays a major role in maintaining equilibrium during rapid
motions and motions with rapidly changing direction through a predictive
function. In this case the cerebellum doesn't wait for proprioceptive signals from
the body as they might be too late. Instead, it uses the proprioceptive signals
from the vestibular apparatus and the periphery to compute the speed and
direction of the present movement, then, it predicts the direction and speed of the
coming movement and responds to correct any disturbance in equilibrium during
this movement.
[II]CONTROL OF MUSCLE TONE
The neocerebellum and paleocerebellum facilitate the muscle tone, whilst the
archicerebellum inhibits it. During rest, the fac ilitatory influence predominates.
Although the facilitatory output originate from the cerebrocerebllum and
spinocerebellum, the spinocerebeilum is the main part of the cerebellum
concerned with controlling the muscle tone through increasing or decreasing the
facilitatory output.

130
THE CEREBELLAR STRETCH REFLEX: This reflex operates when a contracting
muscle meets an unexpected load. Signals from muscle spindles are transmitted
to the spinocerehellum via the spinocerebellar tracts. The spinocerebellum
responds by output signals to the alpha and gamma motor neurons of the spinal
cord to increase the muscle tone to the appropriate required level (the load
reflex).
[III] CONTROL OF VOLUNTARY MOVEMENT
The cerebrocerebellum influences voluntary movement through the
following functions:
1. PLANNING: The cerebrocerebellum receives input signals from the premotor
cortex and inferior olive before the start of movement to give the cerebellum an
"efferenee copy" of the motor plan. This informs the cerebellum about the
"intention
"
to do a movement.
Through connections with the motor cortex the cerebellum "monitors" the flow
of signals from the motor cortex and corrects any deviation from the planned
sequential order.
2. TIMING OF MOVEMENT: The; cerebellum determines the start and
termination of each of the sequential movement which form a complex movement.
Without this function, the person becomes unable to determine when the next
movement should begin. The succeeding movement begins too early or, more
likely, too late leading to decomposition of movement.
3. JOINING OF MOVEMENT: The dentate nucleus shows the "activity pattern
of the next movement at the same time the present movement is occurring. The
cerebrocerebellum is not concerned with what is happening at a given moment
but with what will happen at the next. This leads to rapid succession of the
sequential movements' one after the other to produce a smooth, joined complex
movement.
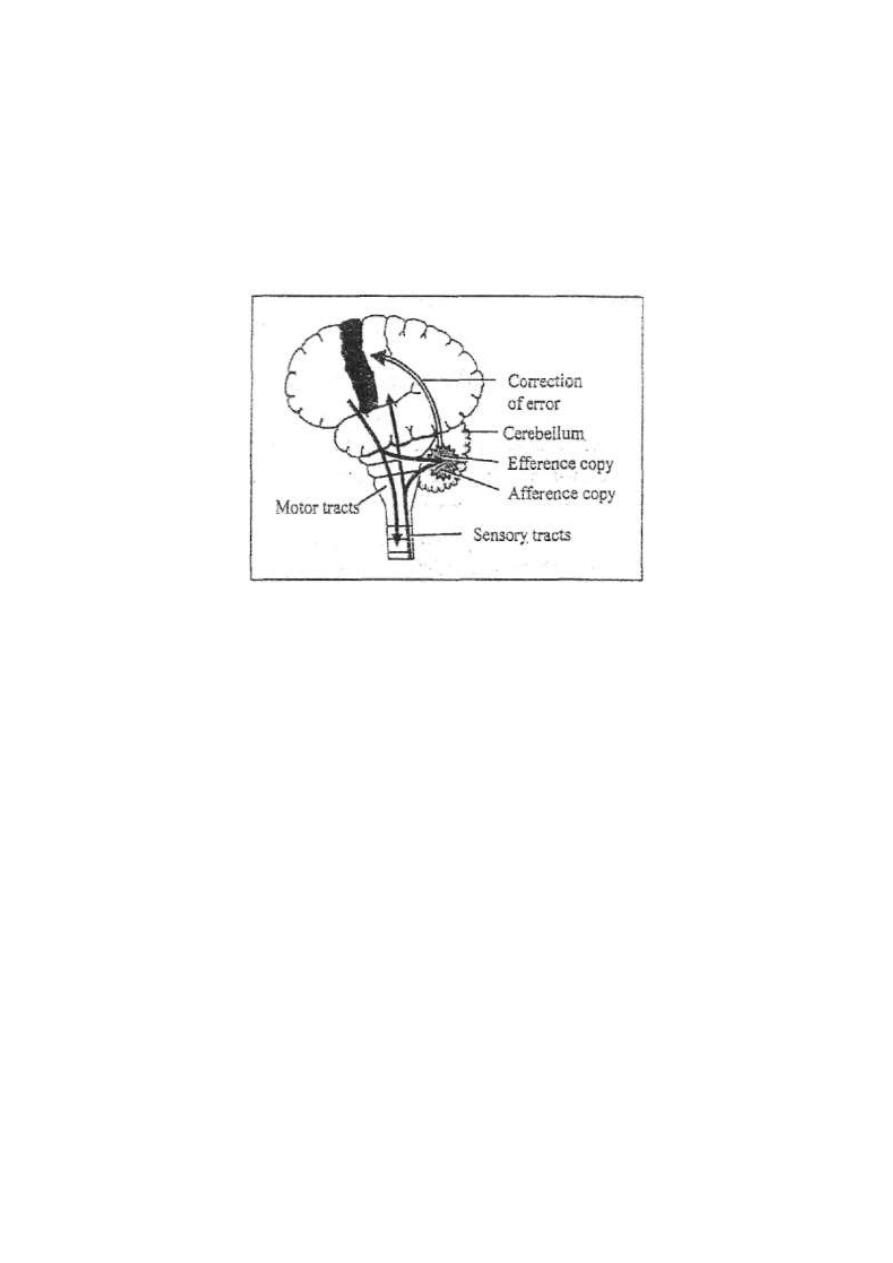
131
4. SERVOCOMPARATOR (force regulator):
The cerebellum (fig. 12-1) receives input signals from:
(i)The premotor cortex and inferior olive, before the start of movement. An
"efference copy" of the plan of movement informs the. cerebellum of the
"intention" to make a movement.
Figure 12-1: The servocomparator function of the cerebellum
(ii) The spinocerebellar tracts, during the movement. An "afference copy" of
the signals reaching the ventral horn cells and proprioceptive signals from the
moving muscles and joints inform the cerebellum about the "performance" of
the movement.
The spinocerebellum then compares the afference with the efference copy; i.e.
the performance with the intention. If there is any error in the performance, the
cerebellum sends correction signals via the descending motor tracts to adjust the
movement to match the intention.
5. DAMPING OF MOVEMENT:
Damping of movement means ending of movement abruptly without oscillation.
The cerebellum acts as a "break" which terminates the movements exactly at the
right time when the desired movement is completed. This is achieved through the
inhibitory output signals from the nuclear cells which occurs in response to input

132
signals in the climbing and mossy fibers.
The absence of this function leads to overshooting of movements and when the
motor cortex corrects the overshooting, the correction overshoots again. This
process of "correction of the overshooting and overshooting of the correction" is
repeated several times before the movement reaches its target. In this way, a
hand or a finger oscillates back and forth past its target before it finally settles on
it. This feature is called "intention tremor" or "kinetic tremor".
6. PREDICTTVE FUNCTION:
As the cerebellum receives sensory input from all sensory organs, it is informed
about the rate and direction of movement. The cerebellum uses this information
to compute and change the rate and direction of movement to reach a certain
target at a certain time; e.g. the thrust of a goal-keeper to catch a kicked football,
or the thrust of a tennis player to meet a coming ball with his racket. In these
cases, visual signals inform the cerebellum about the speed and direction of the
ball movement. Proprioceptive signals inform the cerebellum about the speed and
direction of body movement. The cerebellum adjusts the movement of the body
to meet the ball at a certain time in a certain location.
When a person runs towards a wall, this function enables him to stop just before
the wall without hitting it.
7.
INITIATION AND TERMINATION OF BALLISTIC MOVEMENT:
A ballistic movement is a short movement whose direction and speed is
determined by the initial thrust; e.g. movements of the finger of a typist during typing.
After starting a ballistic movement, its direction and speed cannot be changed.
The rapidity and shortness of movement do not allow the cerebellum to carry out
its servocomparator adjustive function. The role of the cerebellum here is to
potentiate the starting of the movement to allow a rapid onset and to terminate the

133
movement abruptly without overshooting.
Potentiation of the starting is achieved by the initial excitatory output signals of
the mossy fiber circuit. The abrupt termination is achieved by the inhibitory
output signals which follow the excitatory signals.
THE CEREBELLAR SYNDROME
The cerebellar syndrome results from lesions that involve the cerebellar nuclei.
Removal of large parts of the cerebellar cortex without injuring the deep nuclei
leaves little long term effects. That is why neurosurgeons take great care not to
injure the cerebellar nuclei during operations on the cerebellum.
The effects of the cerebellar syndrome appear on the same side of the lesion.
They include three main features; i.e. asthenia, atonia, ataxia (AAA).
1. ASTHENIA: Asthenia means lack of strength. The patient feels muscle
weakness cue to me difficulty in initiating and maintaining muscle contraction.
Asthenia appears because the function of the cerebellum in potentiating motor
signals is lost.
2. ATONIA (or hypotonia): Atonia means lack of muscle tone. This occurs due
to loss of the facilitatory effect of the dentate and interpositus nuclei. Atonia is
manifested by the flaccid feel of the muscles and pendular knee jerk.
3. ATAXIA: Ataxia means in coordination of voluntary movements. Cerebellar
ataxia is manifested by:
(a)
Dysmetria: It means inability to adjust a motor act for a certain distance. The
movement usually overshoots the target (past pointing or hypermetria), or
sometimes stops short of the target (hypometria). Dysmetria is caused by failure
of the timing and damping functions of the cerebellum
.
(b) Kinetic tremor (intention tremor): This is an oscillatory movement which
occurs on doing a voluntary movement. The tremor is absent during rest.
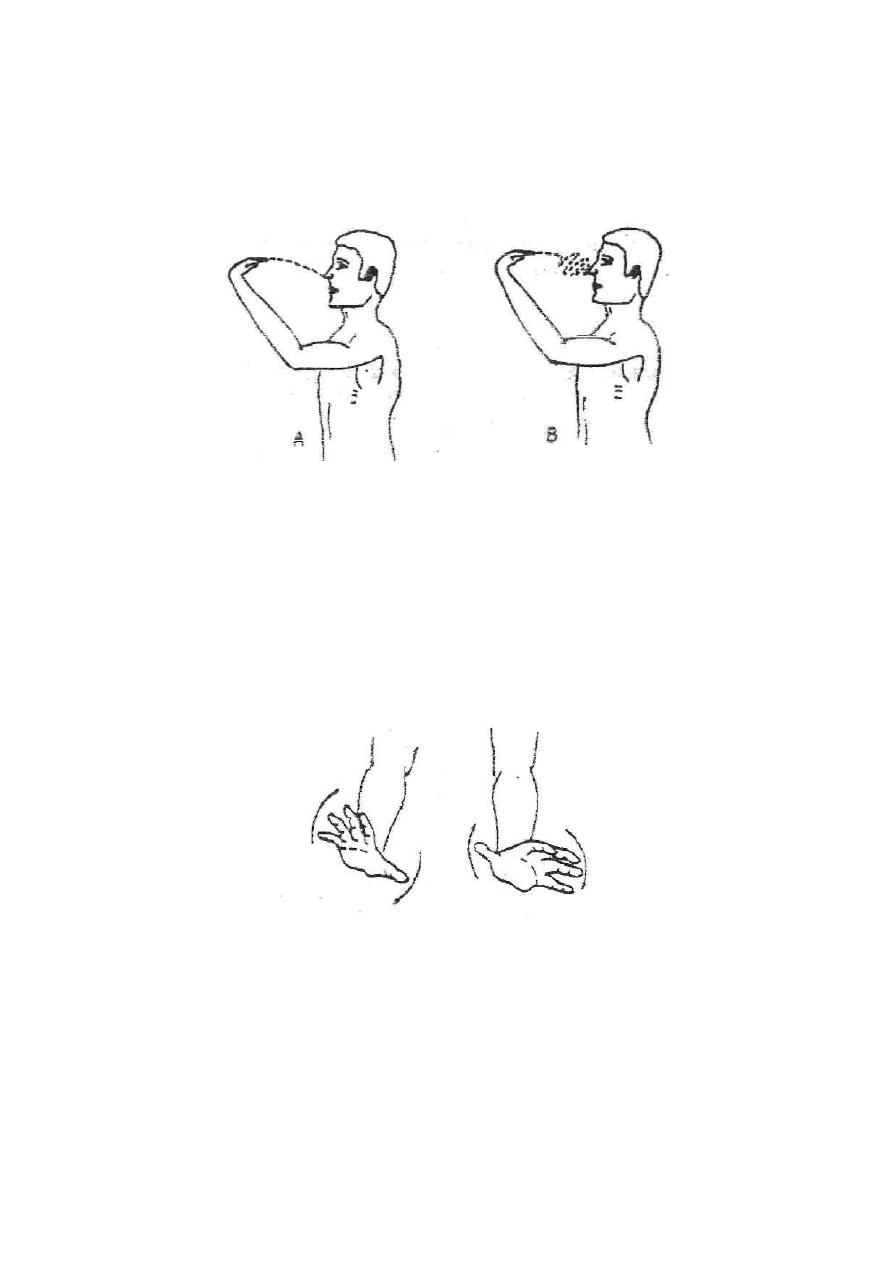
134
The cerebellar kinetic tremor results from a series of "correction of overshooting
and overshooting of the correction". This disability is tested by the finger to-nose
test (fig. 12-3).
Figure 12 - 3: Finger to nose test in a normal person (A) and in a person with
cerebellar syndrome (B).
(c) Adiadokokinesia: This means the inability to do rapid alternating
movements; e.g. pronation and. supination of the hand (fig. 12-4). This is due to
failure of the predictive and joining functions of the cerebellum.
Figure 1 2 -4 : Adiadokokinesia.
(d) Decomposition of movement: Movements which involve more than one
joint are broken up to their component simple movements.
(e) Dysarthria: this means difficulty in producing clear, normal speech. The lack
of predictive and joining functions leads to in coordination of the buccal, laryngeal and
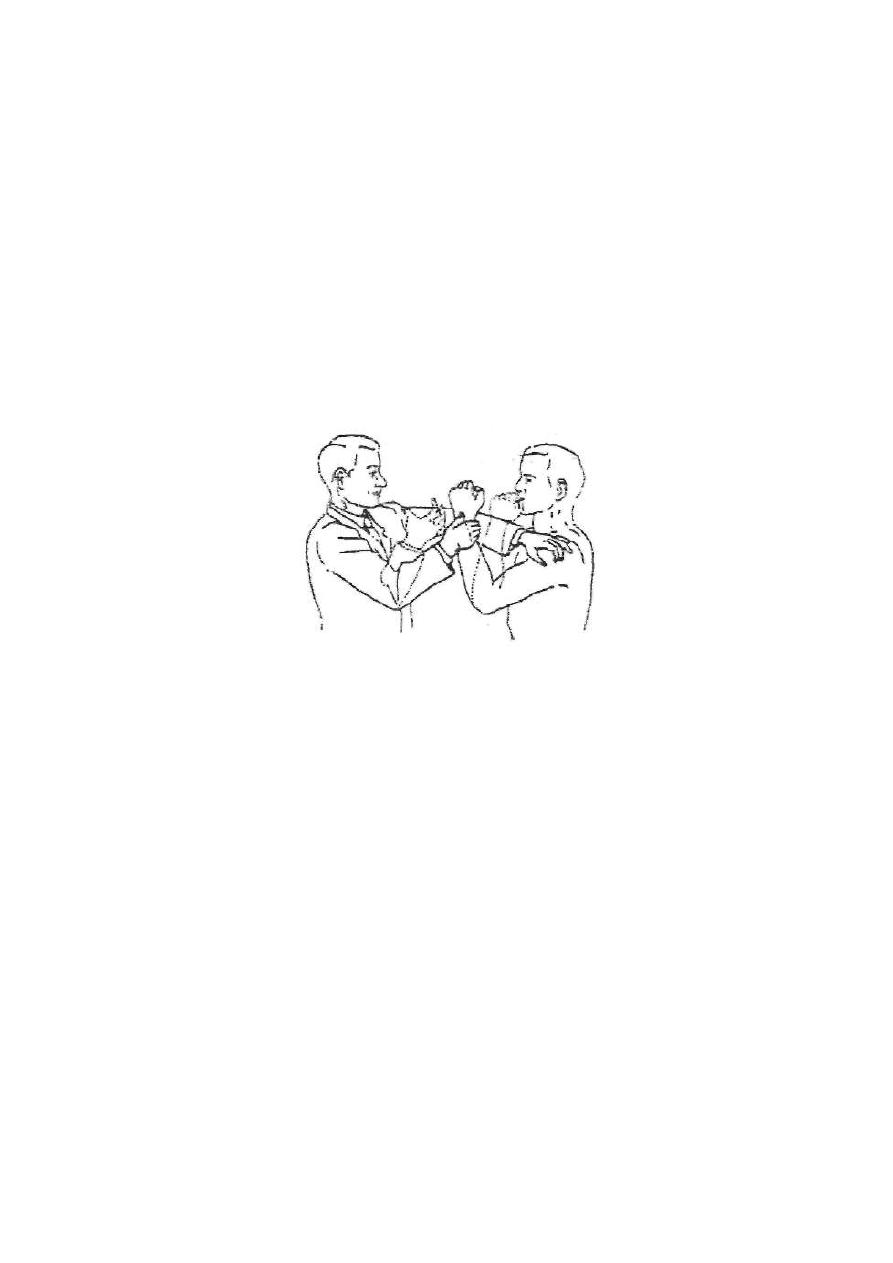
135
respiratory movements. This results in jumbled speech in which some syllables are too
long, others are too short. The volume of sound is poorly controlled; it oscillates
between loud and faint. Sometimes, speech becomes scanning or staccato speech,
where speech is cut off into short syllables separated by regular intervals, e.g. I
re-cei-ved an in-vi-ta-tion.
(f) Rebounding: This is manifested by an overshooting of a limb when
a resistance to its movement is suddenly withdrawn. This disability is
tested by the "arm flexion test" (fig 12-5)
Figure 12 - 5: The arm flexion test
.
(g) Nystagmus: T h is is an oscillatory movement of the eye. Cerebellar
nystagmus appears on lateral gazing. It results from lesions in the
archicerebellum.
(h) Staggering gait: The patient walks on a wide base in a drunken, swaying or
zigzag manner and tends to fall on the diseased side. This is a manifestation of a
lesion in the archicerebellum.
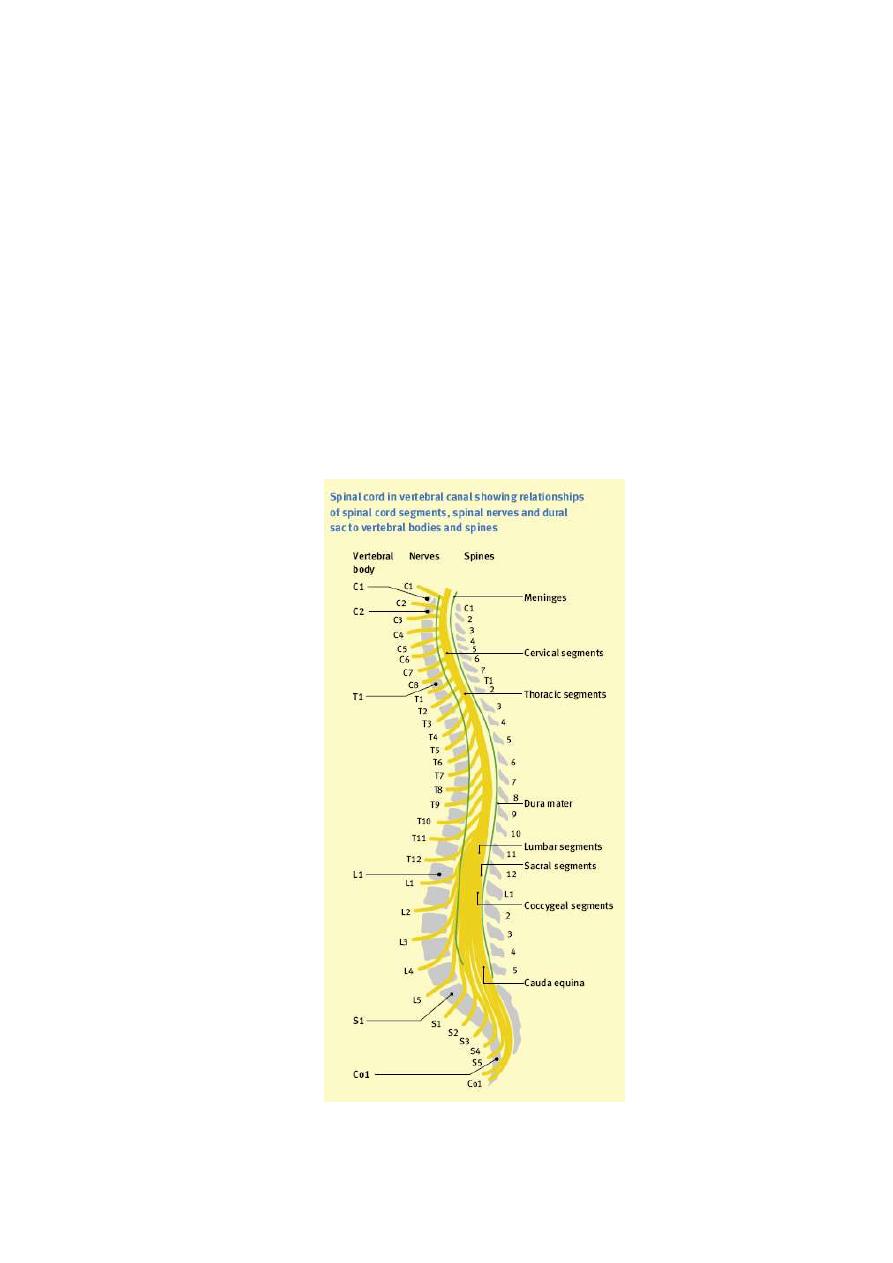
136
PHYSIOLOGY
Dr. Basim Mohamad Alwan Lecture 13
REFLEXES
REFLEX ARC
It is the basic unit of the integrated neural activity.
It consists of sense organ, an afferent neuron, one or more synapses, and an
efferent neuron.
The simplest reflex arc is the one with a single synapse between the afferent
and the efferent neurons such as the "monosynaptic".
Those more than one interneuron are the "polysynaptic".
The number of the synapses in the arcs varying from 2 to many thousands.
Fig. 13-1 Spinal cord segment and vertebral canal

137
ORGANIZATION OF THE SPINAL CORD FOR MOTOR FUNCTIONS (fig. 14-2)
Each segment of the spinal cord between one spinal nerve and the next has
several million neurons in the gray matter.
These neurons are:
Anterior Motor Neurons
Several thousand neurons are located in each segment of the anterior horn of
spinal cord.
They are 50-100% larger than most of others.
They give rise to the nerve fibers that leaves the spinal cord by way of the
ventral roots and innervates the skeletal muscle fibers.
They are of 2 types:
Alpha Motor Neurons
They give rise to type A alpha nerve fibers that innervate the large skeletal
muscle fibers.
Stimulation of a single nerve fiber excites from as few as 3 to as many as
hundred skeletal muscle fibers.
Gamma Motor Neurons
These neurons transmit impulse through type A gamma fibers.
They are much smaller and reach special skeletal fibers called "intrafusal
fibers", which is part of the muscle spindle.
Interneuron
These present in all areas of the gray matter.
They are numerous about 30 times as the anterior motor neurons.
They are small, highly excitable, and capable of firing as rapidly as 1,500
times / second.
They have many interconnections one with the other and many of them
directly innervate the anterior motor neurons.
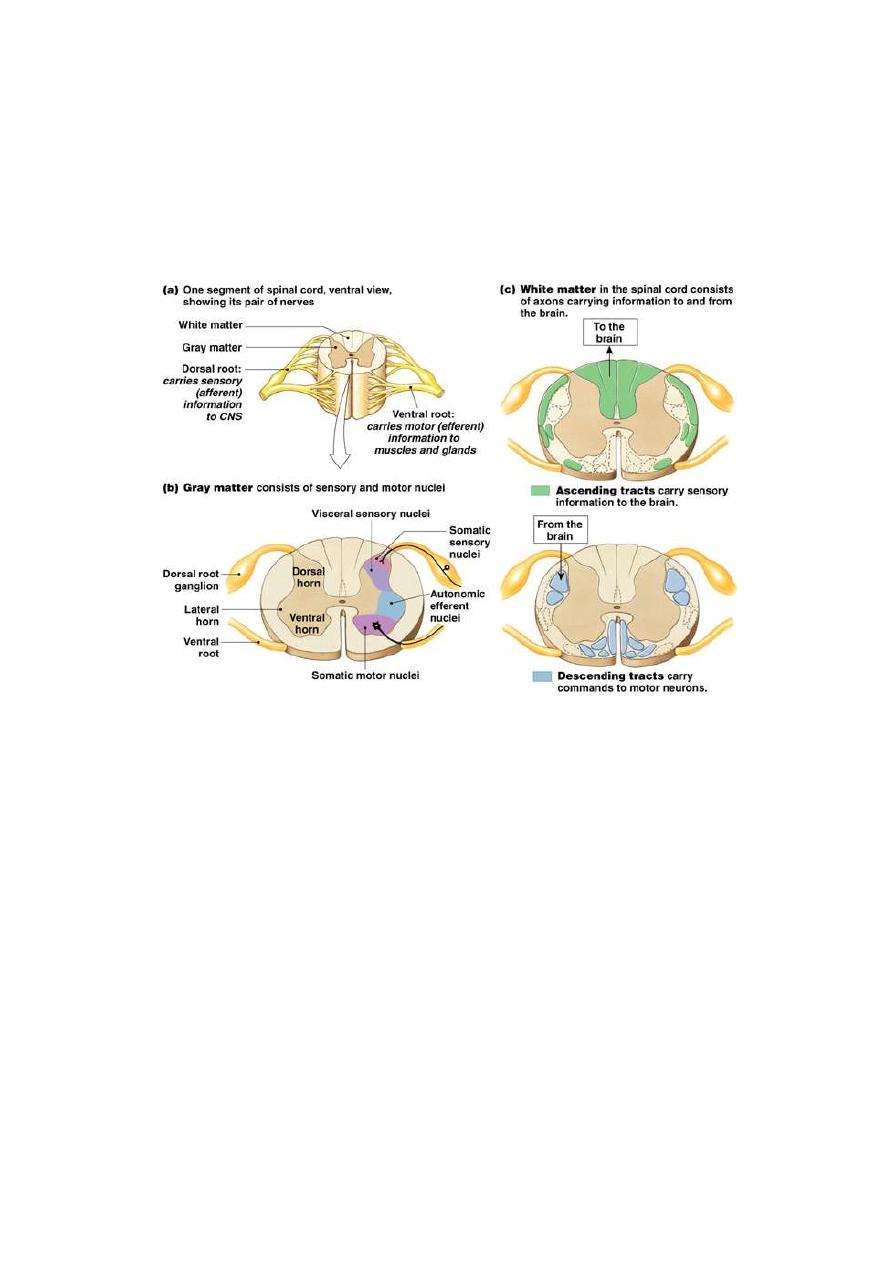
138
Fig. 13-2 Spinal cord organization for sensory, motor and autonomic functions

139
MUSCLE SENSORY RECEPTORS (fig.13-3, 13-4)
Proper control of muscle function requires not only excitation of the muscle by
the anterior motor neurons but also continuous sensory feedback information
from each muscle to the spinal cord giving the status of the muscle at each
instant, that is:
I.
What is the length of the muscle?
II.
What is its tension?
III.
How rapidly is its length or tension changing?
To provide this information, the muscle and its tendon are supplied
abundantly with 2 special types of sensory receptors "muscle spindles and
Golgi tendon organs".
LENGTH-MONITORING SYSTEM AND THE STRETCH REFLEX
Absolute muscle length and changes in muscle length are monitored by
stretch receptors embedded within the muscle.
There are 2 kinds of stretch receptors:
I.
One respond best to how much the muscle has been stretched.
II.
Other responds to both the magnitude of the stretch and the speed with
which it occurs.
These receptors consist of afferent nerve fiber endings that are wrapped
around modified muscle fibers. Several of which are enclosed in a connective
tissue capsule. The entire structure is called "muscle spindle".
Each spindle is 3-10 mm long.
It is build around 3-12 very small modified intrafusal fibers that are pointed at
their ends and attached to glycocalyx of the surrounding large "extrafusal
fiber".
Intrafusal skeletal muscle fibers
They are small muscle fiber
The central region of which has either no or few actin and myosin filaments.
Therefore, this central portion does not contract when the ends do. Instead
they function as sensory receptors.
The end portions that do contract are excited by small gamma motor neurons.
They are of 2 types:
Nuclear-bag fibers
a. They are 1-3 in each spindle.
b. There is large number of nuclei congregated in an expanded bag in the
central portion of the receptors.
c. They are innervated by primary nerve endings

140
Nuclear-chain fibers
a. They are 3-9 in each spindle.
b. They are about 1/2 in diameter and 1/2 as long as the nuclear-bag fibers.
c. Their nuclei aligned in a chain throughout the receptor area.
d. They are innervated by primary and secondary nerve endings.
e. Their ends are connected to the nuclear-bag fibers.
The sensory fibers which originate in the central portion and stimulated by
stretching of this mid portion of the spindle are of 2 types:
The Primary Endings (Annulospiral) Fibers
i.
They are large sensory fibers encircles the very center of the receptor area.
ii.
They are termination of type Ia afferent fibers.
iii.
They have 17 µm diameters.
iv.
They transmit signals to the spinal cord at a velocity of 70-120 m/second.
v.
They discharge most rapidly while the muscle being stretched and less rapidly
during sustained stretch. Thus, respond to changes in length and change in
the rate of stretch.
The Secondary (Flowerspray) Fibers
i.
They are the termination of usually one and some times two smaller type II
sensory nerve fibers.
ii.
Their diameter is 8 µm.
iii.
They innervate the receptor region on one side of the primary ending.
iv.
They discharge at increasing rate through out period when the muscle is
stretched.
v.
They respond to change in the length alone.
Note: Both primary and secondary nerve endings are stimulated when the spindle is
stretched but the pattern of response is different.
Fig. 13-3 muscle spindle
141
Fig. 13-4 muscle spindle
The muscle spindles are parallel to the extrafusal fibers, such that stretch of
the muscle by external force pulls on the intrafusal fibers, stretching them and
activating their receptor endings with reflex contraction of the extrafusal fibers
of the muscle.
The more the muscle is stretched or the faster it stretched, the greater the
rate of the receptor firing.
In contrast, contraction of the extrafusal fibers and the resultant shortening of
the muscle remove tension on the spindle and slow the rate of firing of the
stretch receptor.
When the afferent fibers form the muscle spindle enter the CNS, they divide
into branches that take several different paths:
First Path:
Directly stimulates motor neuron going back to the muscle that was stretched,
thereby completing a reflex arc known as the "stretch reflex".
This reflex is probably the most familiar in the form of the knee jerk.
1. The examiner taps the pattelar tendon which passes over the knee and
connects extensor muscles in the thigh to the tibia in the lower leg.
2. As the tendon is pushed in, and thereby stretched by tapping.
3. The thigh muscles to which it attached are stretched and all the stretched
receptors within these muscles are activated.
4. More action potentials are generated in the afferent nerve fibers from the
stretched receptors.
5. They are transmitted to the motor neurons that control these muscles.
6. The motor units are stimulated
7. The thigh muscles are shortens
8. The patient's lower leg is extended to give the knee jerk.

142
Note: In contrast to the knee jerk, during normal movement, stretch receptors are
rarely all activated at the same time, nor are they activated so strongly.
Moreover, movement occurs in response to the integration of many types of
local descending controls.
Second Path
1. The afferent nerve fibers from stretch receptors end on interneuron.
2. When activated, inhibit the motor neuron controlling the antagonistic muscles
(in the knee jerk, the flexor muscles of the knee are inhibited).
Reciprocal Innervations: The activation of one muscle with simultaneous inhibition
of its antagonistic muscle.
Third Path
1. Afferent nerve fibers activate motor neurons of synergistic muscles, that is,
muscles whose contraction assists the intended motion (in the knee jerk,
other extensor muscles).
2. The muscles activated are on the same side of the body as the receptors and
the response therefore is ipsilateral.
Fourth Path
1. Afferent nerve fibers continue to the brain stem.
2. They synapse there with interneuron which forms the next link in the pathway
that conveys information about the muscle length to areas of the brain dealing
with motor control.
ALPHA-GAMMA COACTIVATION
When the muscle shortens, stretch of the intrafusal fibers decreased, and at this
time, the spindle stretch receptors go completely slack, and they no stop firing action
potentials.
But:
In this situation, there can be no indication of any further changes in the muscle
length.
Physiologically:
To prevent this loss of information, the two ends of each intrafusal muscle fiber are
also stimulated to contract during the shortening of the extrafusal muscle fibers.
Thus, maintaining tension in the central region of the intrafusal fibers, where the
stretch receptors are located.
Note: The intrafusal muscle fibers are not large enough or strong enough to shorten
a whole muscle and move joints; their sole job is to maintain tension on the
spindle stretch receptors.
Note: Both alpha and gamma motor neurons are activated by interneurons in their
immediate vicinity and by neurons of the descending pathways. In other
words, they are coactivated, that is, excited at almost the same time during
many voluntary and involuntary movements.
143
TENSION-MONITORING SYSTEM AND GOLGI TENDON REFLEX
Any given set of inputs to a given set of motor neurons can lead to various
degrees of tension in the muscles they innervate depending on:
a. Muscle length
b. Load on the muscle
c. Degree of muscle fatigue
Therefore, feedback is necessary to inform the motor control systems of the
tension actually achieved.
Such feedback is provided by vision and by Golgi tendon organs which
monitors how much tension is being exerted by the contracting motor units.
Fig. (13-5) Golgi tendon organ
i.
They are encapsulated sensory receptors through which a small bundle of
muscle tendon fibers pass.
ii.
They are located in the tendons near their junction with the muscle.
iii.
About 10-15 muscle fibers are usually connected in series with each Golgi
tendon organ.
iv.
Endings of the afferent nerve fibers are wrapped around collagen bundles
in the tendon.
v.
When he attached extrafusal muscle fibers contract, they pull on the
tendon, which straightens the collagen bundles and distorts the receptor
endings, and activating them.
vi.
Thus, the Golgi tendon organs discharge in response to the tension
generated by the contraction and initiates action potentials that are
transmitted to the CNS via large rapidly conducting type IIb nerve fibers.
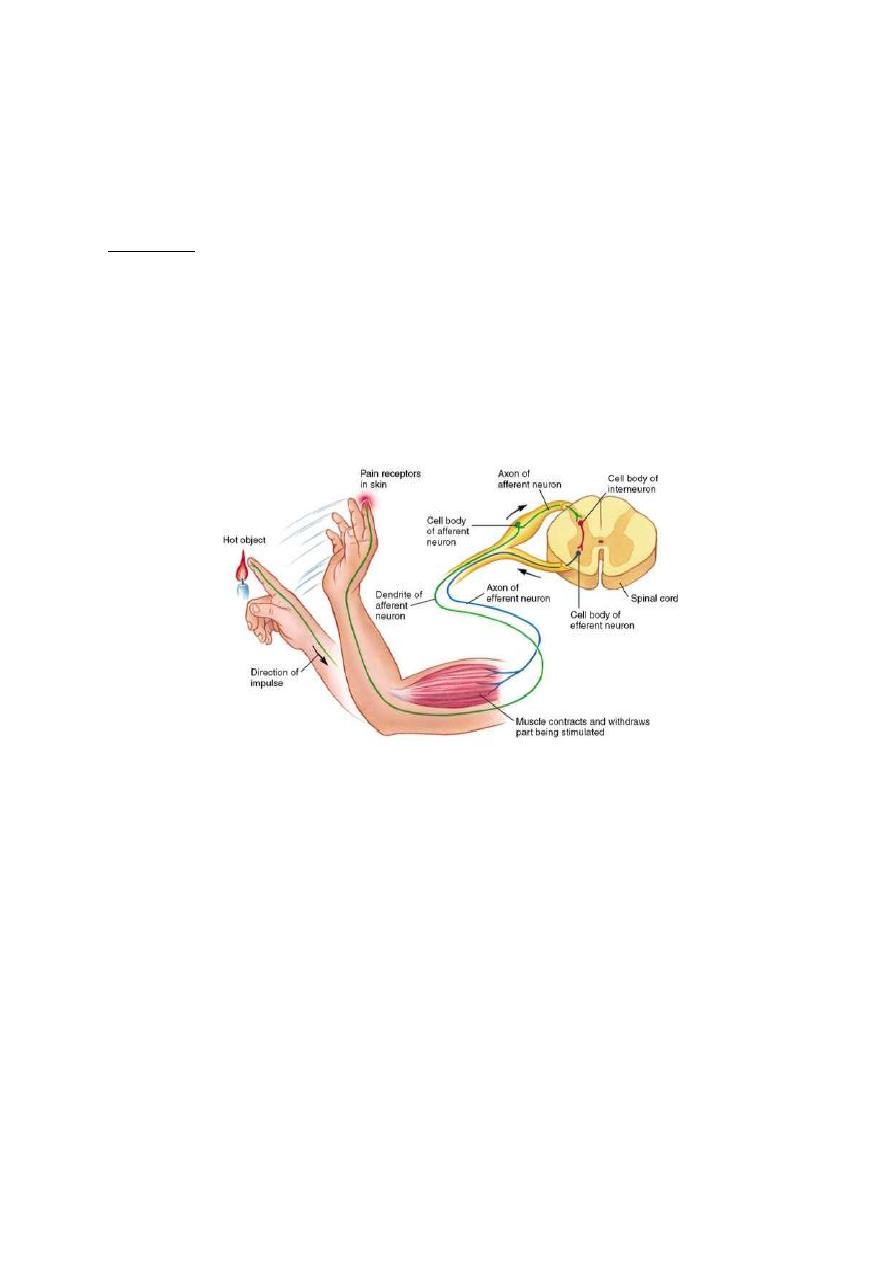
144
vii.
Branches of the afferent neurons cause widespread inhibition via
interneurons, of the motor neurons to the contracting muscle and its
synergists.
viii.
They also stimulate the motor neurons of the antagonistic muscles.
Importance
1. Prevent tearing of the muscle or avulsion of the tendon from its attachments
to the bone.
2. Equalize the contractile forces of the separate muscle fibers (that is fibers
which exert excess tension become inhibited by the reflex and the reverse is
true. This will spread the muscle load over all fibers and would prevent
damage in isolated areas of a muscle where small numbers of fibers might be
overloaded.
Fig. 13-6 Withdrawal reflex
WITHDRAWAL REFLEX (fig.14-6)
Cutaneous sensory stimuli on a limb are likely to cause the flexor muscles of the limb
to contract, thereby stimulating withdrawing the limb from the stimulating object. This
is called "flexor reflex" or the "nociceptive reflex" or simply "pain reflex".
It is polysynaptic reflex
The response includes flexor muscles contraction and inhibition of the
extensor muscles, so that the part stimulated is flexed and withdrawn from the
stimulus.
When a strong stimulus is applied to a limb, the response includes not only
flexion and withdrawal of that limb but also extension of the opposite limb.
This is called "crossed extensor reflex".

145
PHYSIOLOGY
Dr. Basim Mohamad Alwan Lecture 14
SPINAL CORD
Transection of Spinal Cord
(1) Hemi-section (Brown-Séquard Syndrome)
This occurs in only 1/2 of the spinal cord.
The following effects occur:
a) All motor functions are blocked in all segments below the level of transaction.
b) Only some of sensory modalities are lost on the transected side and others
are lost on the other side.
c) Pain, heat, and cold sensations (served by spinothalamic tract) are lost on the
opposite side of the body in all dermatomes 2-6 segments below the level of
transaction.
d) Kinesthetic position, vibration, discrete localization and the two-point
discrimination sensations (served by dorsal and dorsolateral columns) are lost
on side of transaction in all dermatomes below the level of transaction.
e) Discrete light touch sensation is impaired on the side of transaction.
f) Crude touch sensation which is poorly localized still persist because of
transmission in the opposite spinothalamic tract.
146
Fig. 14-1 Brown sequard syndrom
(2) Complete Transverse Section
When the spinal cord is transected in the upper neck, all cord functions including
cord reflexes become depressed and lost. This reaction is called "spinal shock".
Mechanism:
The normal activity of spinal cord neurons depends to a great extent on the
continual tonic excitation by discharges of nerve fibers entering the cord from
higher centers (particularly discharges transmitted through reticulospinal,
vestibulospinal, and corticospinal tracts).
After few hours to few weeks, the spinal neurons gradually regain their
excitability (this is a natural characteristic of the neurons anywhere in the
CNS).
In most non-primates, excitability of cord centers returns essentially to normal
within a few hours to day or so.
In human being, return is delayed fro several weeks and occasionally never
complete. Sometimes, recovery is excessive with resultant hyper excitability
of some or all cord functions.
The following are some of spinal functions specifically affected during or after
spinal shock:
(1) Arterial blood pressure falls to as low as 40 mmHg (blocking of the
sympathetic activity). This returns to normal within a few days.

147
(2) All skeletal muscle reflexes integrated in spinal cord are blocked during initial
stage of shock. Two weeks to several months are needed by those reflexes
to return to normal.
Note: Sometimes, the reflexes become hyper excitable (especially if few facilitatory
pathways remain intact between the brain and the cord whiles the remainder
of the spinal cord is transected.
Note: The first reflex to return normal are stretch reflexes and remnants of steeping
reflexes. In some patients, the knee jerk is returned first and in other patients,
slight contraction of leg reflexes and adductors in response to noxious stimuli.
(3) Sacral reflexes for control of UB and colon evaluation are suppressed for the
first few weeks after cord transaction but then eventually return.
Complication of Cord Transection
I.
They develop negative nitrogen balance and catabolize large amounts of
body proteins.
II.
Decubitus ulcers due to compression of skin circulation by the body
weight.
III.
Hypercalcemia that lead to hypercalciuria and calcium stones.
IV.
The stones in combination with UB paralysis cause urine stasis which
predispose to urinary infection.
V.
Death from septicemia and uremia.
Responses in Chronic Spinal Humans
In chronically quadriplegic humans, there is:
A. Threshold of withdrawal reflex is low (minor noxious stimuli may cause not
only prolonged withdrawal of one extremity but marked flexion-extension
patterns in the other 3 limbs).
B. Repeated flexion movements occur for prolonged periods and contractures of
flexor muscles develop.
C. Stretch reflexes are hyperactive.
D. Reflex contractions of the full UB and rectum occur. The hyperactive UB
reflexes keep it in a shrunken state leading to hypertrophy and fibrosis of its
wall.
E. Blood pressure is generally normal at rest, but wide swings in the pressure
are common as the feedback regulation by baroreceptor reflexes is absent.
F. Bouts of sweating and blanching of the skin.
G. Genital manipulation in males produces erection and even ejaculation.
H. The Mass reflex (strong nociceptive stimulus to skin or excessive filling of a
viscous (over distention of the UB or gut) can cause massive discharge of
large portions of the cord. The effects are:
Major part of the body goes into strong flexor spasm

148
The colon and UB are likely to evacuate
The blood pressure rises to a systolic pressure of > 200 mmHg
Profuse sweating of large areas of the body.
DECEREBRATE RIGIDITY
Sectioning brainstem below the midlevel of mesencephalon (leaving both
pontine and medullary reticular system as well as vestibular system intact)
leads the animal to develop "decerebrate rigidity".
The rigidity occurs in antigravity muscles (the muscles of the neck, trunk and
extensors of the leg).
Mechanism
Blocking of normal strong excitatory input to medullary reticular nuclei from cerebral
cortex, red nucleus and basal ganglia → causing the medullary reticular inhibitory
system become nonfunctional → allowing full activity of the pontine excitatory
system and development of rigidity.
The antigravity muscles exhibit phenomenon called spaticity as well as rigidity (this
means any attempt to change position of a limb or other parts of the body, especially
attempts to stretch the muscles suddenly, is resisted by powerful stretch reflex).
This is because:
The pontine and vestibular antigravity signals to the cord selectively excite gamma
moor neurons in the spinal cord much more than they excite α motor neurons. This is
tightens the intrafusal muscle fibers if the muscle spindles which in turn strongly
sensitizes the stretch reflex feedback loop.
Tonic labyrinthine Reflexes
The decerebrate animal stays in the position in which it put; no righting
responses are present, like:
1. Placing the animal on its back causes maximal extension of the four limbs.
2. Turning the animal to one side causes the rigidity to decreases
3. Putting the animal in prone position causes minimal rigidity
These changes in rigidity are initiated by the action of gravity on otolith organs
and affected via vestibulospinal tracts.
Tonic Neck Reflexes
Moving the head of decerebrate animal relative to the body produces changes
in the pattern of gravity.
1. Turning the head to one side causes the limbs on that side become rigidly
extended while contralateral limbs become less so.

149
2. Flexion of the head causes flexion of the forelimbs and continued extension of
hindlimbs.
3. Extension of the head causes flexion of the hindlimbs and extension of the
forelimbs.
These reflexes are initiated by stretch of proprioceptors in the upper part of
the neck and they can be sustained for long periods of time.
Upper and Lower Motor Neurons
(1) Lesions of the lower motor neurons (spinal and cranial motor neurons that
directly innervate the muscles) are associated with flaccid paralysis muscular
atrophy and absence of the reflex responses.
(2) Upper motor neuron lesions (the neurons in the brain and spinal cord that
activate the motor neurons are destroyed) are associated with spastic
paralysis and hyperactive stretch reflexes in the absence of the muscle
atrophy. It is of 3 types:
a. Lesions in the extrapyramidal posture regulating pathway cause spastic
paralysis.
b. Lesions limited to the pyramidal tracts causes weakness (paresis rather than
paralysis) and the affected musculature is generally hypotonic.
c. Cerebellar lesions cause incoordination.
150
Physiology
Dr. Basim Mohamad Alwan Lecture 15
THE HYPOTHALAMUS
The hypothalamus (fig. 15-1) is the part of the diencephalon which forms the
floor and part of the lateral wall of the third ventricle. It is a small structure that
weighs about 5 gm, but is the most important part of the brain in controlling
homeostasis. It is connected to the pituitary gland by the pituitary stalk
(hypophysial stalk). The hypothalamus is a major central component of the
limbic system.
Figure 15 - 1: The medial surface of the right hypothalamus
151
THE HYPOTHALAMIC NUCLEI
The hypothalamic nuclei (fig. 15-l & 15-2) are classified into five groups; anterior,
middle, posterior, lateral and periventricular.
1. The anterior group: Which includes the anterior, the preoptic, the supraoptic,
the paraventricular, and the suprachiasmaric nuclei?
2. The middle group: Which includes the arcuate, the ventromedial and dorsal
nuclei.
3. The posterior group: Which includes the posterior nucleus, the premamillary
nucleus, and medial and lateral mamillary nuclei?
4. The lateral nucleus: Which forms one large nuclear mass?
5. The periventricular nucleus: Which is a thin sheet of gray matter adjacent to the
third ventricle?
Figure 15-2: Coronal (frontal plane) section in the hypothalamus showing the
lateral, periventricular, and paraventricular nuclei.

152
NERVOUS CONNECTIONS OF THE HYPOTHALAMUS
The hypothalamus receives input signals from:
1. The limbic system; the limbic lobe of the cerebral cortex (through the medial
forebrain bundle), the amygdala and the hippocampus.
2. The reticular formation of the brainstem; the sensory reticular formation,
the nucleus ceruleus, the raphe nuclei, and the nucleus of the tractus solitarius.
3. The retina, through fibers in the optic nerve which terminate in the
suprachiasmatic nucleus.
4. The thalamus.
The hypothalamus sends output fibers in three directions:
1. Upwards: To the thalamus and limbic system (the limbic lobe, the amygdala and
the hippocampus).
2. Downwards: To the reticular formation of the brainstem. Through the reticular
formation, the hypothalamus is connected to the vital centers in the brainstem and
spinal cord which control the autonomic functions.
3. To the posterior pituitary gland: through the hypothalamo hypophysial tract.
FUNCTIONS OF THE HYPOTHALAMUS
[I] AUTONOMIC FUNCTION
The lateral, dorsomedial; and posterior nuclei of the hypothalamus are considered as
a "Sympathetic center" which responds to emotions by generalized sympathetic
activation (the alarm response).
'The sympathetic center includes adrenaline and noradrenalin secretion centers
which selectively control the secretion of these catecholamine's from the adrenal
medulla in different conditions.
Stimulation of certain parts of the hypothalamus produces parasympathetic

153
effects; e.g. micturition. But the control of the parasympathetic functions by the
hypothalamus is very limited and there is "no parasympathetic center' as such in
the hypothalamus.
[II] ENDOCRINAL FUNCTION
The hypothalamus controls the secretion of hormones from the anterior and
posterior pituitary glands.
CONTROL OF SECRETION OF THE ANTERIOR PITUITARY GLAND
Some nuclei in the hypothalamus contain neurosecretory cells which produce
releasing or inhibiting hormones. These hormones stimulate or inhibit the release of
hormones from the anterior pituitary gland. The secretory hypothalamic cells are
grouped together in an area called "the hypophysiotropic area". This area includes
several nuclei; i.e. the paraventricular, the arcuate, and the ventromedial nuclei. The
axons of the neurosecretory hypophysiotropic cells terminate in the median
eminence, which is the basal part of the hypothalamus, just behind the optic chiasma.
The releasing and inhibiting hormones move inside the hypothalamic neurons by
axoplasmic flow to their terminals in the median eminence where they are released.
From the median eminence, they reach the anterior pituitary via the blood in the
hypothalamo-hypophysial portal circulation.
154
Table 15 -1: The hypothalamic releasing and inhibiting hormones.
Hypothalamic hormone
Main site of formation
The affected ant. Pit. hormone
l.TRH
(Thyrotropin-Releasing Hormone)
Paraventricular area
TSH (Thyrotropin, also called
Thyroid Stimulating
Hormone)
2.CRH
(Corticotropin-Releasing Hormone)
Paraventricular area
ACTH
(Adrenocorticotrophic Hormone)
3.GnRH
(Gonadotropin-Releasmg Hormone)
Arcuate nucleus
Follicle-Stimulating Hormone
and Leutinizing Hormone (both are
called pituitary Gonadotropins)
4.PIHand PRH
(Prolactm-Inhibiting; Hormone and
Prolactin-Releasing Hormone)
Arcuate nucleus
Prolactin
5. GRH and GIH
Growth hormone-Releasing Hormone and
Growth hormone- Inhibiting Hormone.
Ventromedial
nucleus and nearby areas
GH
Growth Hormone (also called
somatotropin or somatotrophic
hormone -STH)
CONTROL OF SECRETION OF THE POSTERIOR PITUITARY GLAND
The supraoptic and paraventricular nuclei of the -hypothalamus are connected to the
posterior pituitary through the hypothalamo-hypophysial tract. Two types of
specific neurosecretory cells in these nuclei synthesize the hormones oxytocin and
antidiuretic hormone (ADH). These hormones flow down in the axoplasm of the
neurosecretory cells to the axon terminals in the posterior pituitary where they
remain stored in secretary vesicles inside the nerve terminals. The hormones are
released on the arrival of the proper nerve signals from the supraoptic and
paraventricular nuclei. The hormones then diffuse through the walls of the
capillaries of the posterior pituitary into the blood stream.

155
[III] THERMOREGULATION
The hypothalamic thermostat consists of a "heat loss center" in the anterior
"hypothalamus and a "heat gain center" in the posterior hypothalamus. Both
centers are interconnected and work in a reciprocal manner.
Stimulation of the heat loss center leads to cutaneous vasodilation and profuse
sweating. Its damage leads to high liability to hyperthermia. Stimulation of the heat
gain center leads to cutaneous vasoconstriction, increased muscle tone, shivering
and stimulation of catecholamine secretion. Its damage leads to high liability to
hypothermia.
The heat loss center contains thermosensitive cells. These cells are sensitive to any
change in blood temperature (± 0.02°C) which is an indicator of the body core
temperature. It responds mainly to any increase in core temperature to prevent
hyperthermia.
The heat gain center contains "thermoresponsive" cells which are not
thermosensitive. These cells respond to input signals coming from
cutaneous thermoreceptors which monitor the surface temperature. It responds
mainly to cooling of the skin to prevent hypothermia.
[IV] CONTROL OF FOOD INTAKE
The hypothalamus contains a food intake controlling system "hypothalamic
appestat" which controls the appetite for food. It consists of two centers; a
"feeding center" in the lateral hypothalamus which stimulates the appetite, and a
"satiety center" in the medial hypothalamus (ventromedial nucleus) which inhibits
the appetite. The satiety center acts by inhibiting the inherent tonic activity of the
feeding center. The damage of the satiety center leads to hyperphagia. The animal
eats large amount of any available edible food, thus leading to obesity
(hypothalamic obesity) (fig. 15-3). Damage of the feeding center leads to severe
anorexia which could be fatal
The hypothalamic appestat is adjusted to a "set point" to maintain a specific body

156
weight for each individual. If a person is starved for some time, then left free to eat
afterwards, he eats only enough to restore his prestarvation weight. Also if a
person is overfed for some time and left free to eat afterwards, he eats too little until
he regains his previous weight.
The amygdaloid nucleus is closely associated with feeding. It is concerned with
"sorting out" of food into edible or inedible. Lesion in the amygdaloid nuclei leads
to hyperphagia, but unlike animals with lesions in the hypothalamic satiety center,
the animals with amygdaloid lesions try to eat any available object
(omniphagia). They eat some of any available object before they discover if it is
edible or not (oral exploration).
Figure 15-3: The effects of a lesion in the hypothalamic satiety center (left)
and in the feeding center (right).
[V] CONTROL OF WATER BALANCE
Thirst center is found in the lateral hypothalamus. It receives input signals from
different sources:
1. From osmoreceptors in the anterior hypothalamus.
2. From osmoreceptors in the liver.
3. From volume receptors in the right atrium and large veins.

157
4. From subfornical organ (SFO) and organum vasculosum of lamina
terminalis (OVLT). These organs are circumventricular organs found at the
anteroventral border of the third ventricle (AV3V) outside the blood-brain barrier.
They are stimulated by angiotensin II (AH) which is released in cases of
hypovolemia
5. From the limbic system and cerebral cortex;. Psychic and emotional stimuli
can produce or modify thirst sensation.
6. From the mouth and pharynx.
Hypertonicity of the plasma or hypovolemia stimulate thirst center and ADH release
(i.e.: stimulate water intake and" inhibit water loss). Hypotonicity or hypervolemia
has the opposite effect.
[VI] CONTROL OF SALT APPETITE
There is a "salt appetite center" in the anterior hypothalamus very close to the
osmoreceptors. Its cells are sensitive to changes in plasma osmolality, as well as the
level of sodium in the plasma (osmosodium receptors). They are stimulated by
hyponatremia, hypo tonicity or hypovolemia to increase the appetite for salt (craving
for salt).
[VII] CONTROL OF CYCLICAL PHENOMENA
Circadian rhythm occurs in more than 100 parameters of human organs and
functions. E.g. body temperature is lowest in the early morning and highest in the
evening, so is the heart rate. CRH, ACTH and cortisol secretions are highest at 8 AM
and lowest at midnight." Melatonin secretion from the pineal gland increases by
night and decreases in daylight. The menstrual cycle in adult females is an example
of a physiological monthly rhythrn.
The pacemakers of the circadian rhythm are found in two regions of the
hypothalamus.

158
First, is the suprachiasmatic nucleus (SCN) which is connected to the retina by
the retinohypothalamic tract. This tract synchronizes the activity of the SCN to the
circadian light/dark cycle. This nucleus is responsible mainly for regulating the
circadian waking/sleeping rhythm.
Second, is the ventromedial nucleus of the hypothalamus (VMNH) which is
connected with the SCN? It is responsible mainly for temperature, endocrinal and
feeding rhythms.
[VIII] ROLE IN LEARNING AND MEMORY
The hypothalamus contains a reward center in the lateral and ventromedial
nuclei. When stimulated, it gives a sense of reward; i.e. relaxation, pleasure and
satisfaction. There is a punishment center in the periventricular nuclei, when
stimulated it gives a sense of punishment; displeasure fear and terror. These centers
constitute important components of the reward and punishment systems which are
very important for emotions, motivation, memory and learning.
To keep any experience in the long term memory, it needs either of two
mechanisms; to stimulate the reward and punishment system or to be practiced
repeatedly. Otherwise, the experience is easily forgotten and will not add to the
memory stores of the individual.
The hypothalamus is also a relay station in the Papez circuit which is concerned with
short term memory
[IX] CONTROL OF MOTOR RESPONSES TO EMOTIONS
The motor responses to emotions are controlled by complex mechanisms that
involve the association areas of the cerebral cortex, the limbic system and the
hypothalamus. However, stimulation or damage of some hypothalamic areas
produces certain emotions with specific motor responses as follows:
159
FEAR
Fear is an unpleasant emotional state which involves a sense of insecurity because
of impending danger or evil. It is produced by stimulation of the fear center in the
periventricular nuclei. Reactions to fear include cowering, avoidance, and sweating,
pupillary dilation, turning the head from side to side to seek escape and flee.
Amygdaloid nuclei in the temporal lobes activate the fear center in the
hypothalamus.
Bilateral temporal lobectomy abolishes fear sensation. In this case, the subject cannot
evaluate dangers and proceeds towards them without precautions; e.g. handling
dangerous animals as scorpions or snakes, or crossing roads full of rapidly going
vehicles.
RAGE
Rage is violent anger. It is produced by stimulation of a certain area of the lateral
hypothalamus (rage area). This area is tonically inhibited by the ventromedial
nucleus (placidity area), and the limbic association area of the cerebral cortex.
Lesions in the placidity area or the limbic association area produce rage.
Reactions in rage include taking the attack position, generalized sympathetic
stimulation and fighting. In cats there is hissing, spitting, growling and well
directed biting and clawing.
PLACIDITY
Placidity means calmness with little or no response to provocation. It is produced by
stimulation of the ventromedial nucleus of the hypothalamus (the placidity area),
or bilateral damage of the amygdaloid nuclei.
The amygdaloid nuclei facilitate the rage area and inhibit the placidity area or
bilateral amygdaloid damage of the amygdaloid nuclei. The amygdaloid facilitates
the rage area and inhibits the placidity area. Bilateral amygdaloid lesions would then
inhibit the rage area and facilitate the placidity area. If the placidity area is

160
subsequently damaged, placidity changes into rage because the rage area would be
released from the inhibitory influence of the placidity area.
Bilateral destruction of the amygdaloid nuclei was made in Japan on agitated,
violent, aggressive mental patients. The patients turned placid and manageable,
without any sign of hyper sexuality.
[X] SEXUAL BEHAVIOR
Libido and sexual activity is mainly under the control of the cerebral cortex and
limbic system which are sensitive to sex hormones. However, hypothalamus is
involved in the following way:
1. The hypophysiotropic area controls the release of the pituitary gonadotropins,
which in turn control the release of sex hormones from the gonads.
2. The anterior hypothalamus in the female contains estrogen sensitive neurons. When
stimulated, these neurons increase the sexual desire and initiate the heat of sexual
behavior. The female seeks out the male (the enticement reaction). Lesions in this area
abolish this behavior.
3. Stimulation of parts of the lateral hypothalamus produces sexual excitement and
penile erection in male monkeys.
[XI] RELATION TO SLEEP
Fibers of the ascending reticular activating system (ARAS) pass through the
hypothalamus in its way to the thalamus then to the cerebral cortex. ARAS fibers
which desynchronize the cerebral cortex and cause wakefulness pass through the
posterior hypothalamus. So, a lesion in the posterior hypothalamus produces sleep
due to damage of these desynchronizing fibers. An irritative lesion may stimulate
these fibers leading to insomnia

161
EFFECTS OF LESIONS IN THE HYPOTHALAMUS
Lesions in the hypothalamus produce widely variable effects depending on the
locality, the extent and the mode of onset; i.e. rapid (acute) or gradual (chronic). A
lesion may produce one or more of the following manifestations.
1. Endocrinal disturbances:
The most common endocrinal disturbance of hypothalamic origin is diabetes
insipidus which is produced by lesions in the supraoptic "or paraventricular nuclei.
There is lack of ADH secretion from the posterior pituitary. This leads to marked
polyuria and polydipsia.
Another form of endocrinal disturbance is precocious puberty i.e. very early
onset of puberty.
Irregularities of the menstrual cycle occur with lesions in the preoptic area or with
psychic disturbances.
Hyper or hypo function of other glands due to hypothalamic causes are possible but
very rare.
2. Hyperthermia or hypothermia:
Produced by lesions in the hypothalamic thermostat.
3. Hyperphagia or anorexia:
Produced by lesions in the hypothalamic appestat. This leads to either
hypothalamic obesity or severe emaciation.
4. Sleep disturbances (insomnia or hypersomnia):
Destructive lesions in the dorsal hypothalamus produce narcolepsy; i.e. attacks of
strong, irresistible desire to sleep. Sleep lasts for short periods could be only few
minutes of which the patient wakes up fully recovered. Insomnia results from
irritative lesions in the dorsal hypothalamus.

162
5. Disturbances in memory and learning ability:
This is due to interruption of the Papez circuits of the limbic system which are
concerned with short term memory. Lesions in the punishment or reward centers
depress the ability to develop long term memory for important events.
6. Autonomic disturbances:
E.g. Lack of response to emergency situations. Increase or decrease in
catecholamine release, excessive sweating, spontaneous vasodilation (hot flushes) or
vasoconstriction.
7. Emotional disturbances:
E.g. fear, rage, or placidity due to irritation or damage of different hypothalamic
regions.
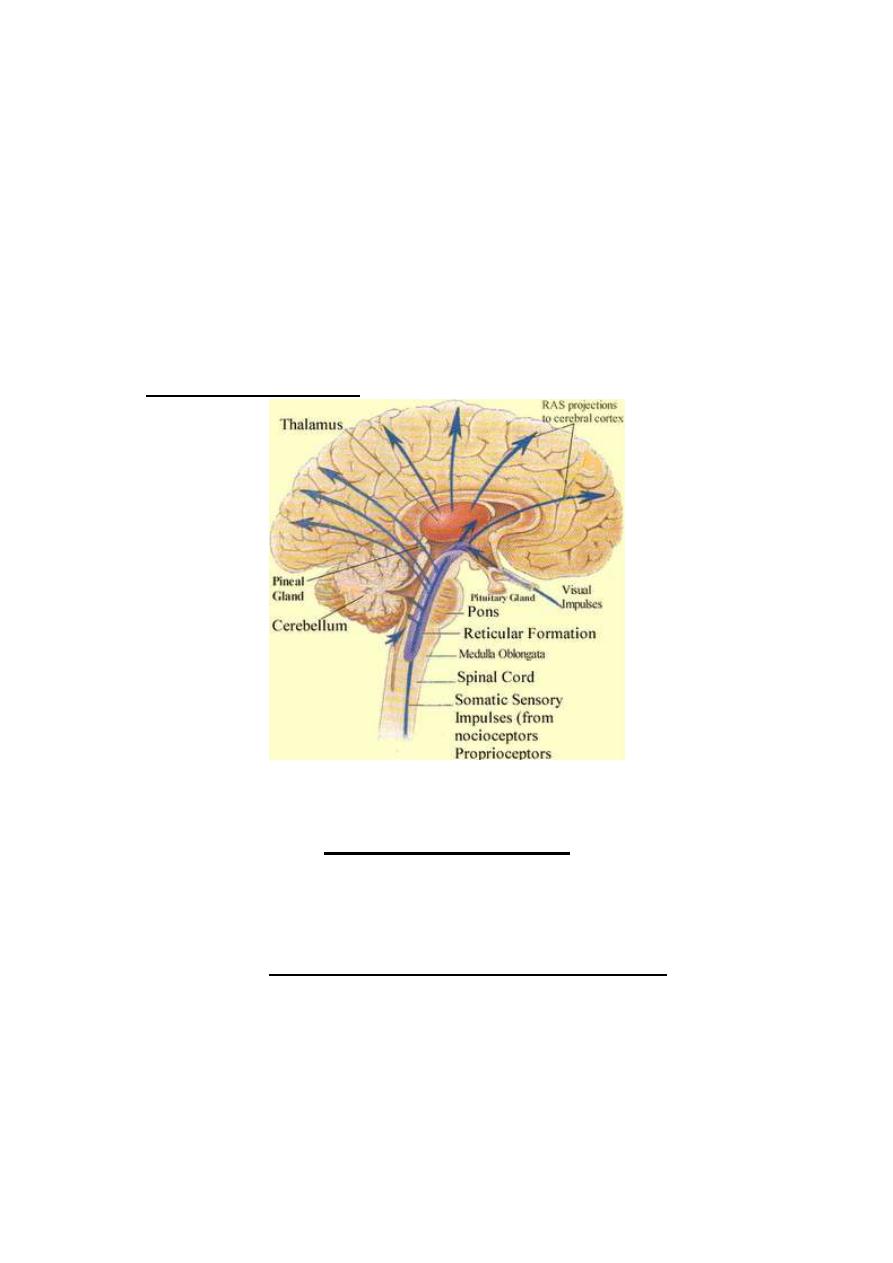
163
PHYSIOLOGY
Dr. Basim Mohamad Alwan Lecture 16
RETICULAR FORMATION
This is a network of neurons located in the brain stem, extending upwards to the
diencephalon (thalamus, hypothalamus and sub thalamus) and downwards to the
upper part of the spinal cord, where it merges with its interneuron's (fig 16-1).
Many nuclei and centers are present within its meshes (respiratory & cardiac
centers, the substantia nigra, the red, vestibular and raphe muclei). It is divided
into sensory and motor parts.
Fig. 16-1 the reticular formation
(A) THE SENSORY PART
This part consists of a large number of small neurons that have multiple
interconnection with each other (which allows marked convergence, divergence
and after discharge).
It receives a rich polysensory input (afferent fibers) from
A. All ascending lemnisci.
B. The visual, auditory and olfactory nervous pathways.
C. The basal ganglia.
D. The cerebellum.
E. The cerebral cortex (corticofugal fibres )
F. The hypothalamus.
G. The vestibular apparatus.

164
(B) THE MOTOR PART
This part consists of large neurons, which receive signals from the sensory
part, and their axons constitute the output (efferent) fibres from the
reticular formation , it contains facilitatory and inhibitory parts .
1. Facilitatory (excitatory) reticular formation: This is located in the
pons and midbrain (specially the former). it has an inherent activity and
the axons of its neurons divide into 2 branches :
a. An ascending branch, which excites the cerebral cortex, and is
called the ascending reticular activating system or ARAS.
b. An descending branch (ventral reticulospinal tract) which
facilitates the spinal centers.
2. Inhibitory reticular formation: this is located mainly in the medulla
oblongata, it has no inherent activity, and its axons descend as the
lateral reticulospinal tract, which inhibits the spinal centers.
FUNCTIONS OF THE RETICULAR FORMATION
1. Control of the level of consciousness via the ascending reticular
activating system.
2. Regulation of the stretch reflex and muscle tone via the
reticulospinal tracts
3. Pain inhibition by the raphe Magnus nucleus.
4. Control of sleep by 2 specific centers.
5. Control of visceral functions (cardiac activity) by controlling the
spinal lateral horn cells.
ASCENDING RETICULAR ACTIVATING SYSTEM
(ARAS)
This is a multineuronal polysynaptic system of nerve fibers that originate at the
facilitatory reticular formation. Its fibers extend upwards, then some project
directly to the cerebral cortex, while the majority relay first at the nonspecific
thalamic nuclei, from which other fibers arise and project diffusely to almost all
parts of the cerebral cortex, the latter pathway is called the reticulo thalamo
cortical pathway.
Functions of the ARAS
The ARAS controls the electric activity of the cerebral cortex, and is concerned
with consciousness and production of the alert response, so reduction of its
activity leads to sleep.

165
FACTORS THAT AFFECT THE ACTIVITY OF
THE ARAS
A. Factors that increase the ARAS activity
1. Sensory signals (especially pain).
2. Signals from the cerebral cortex (via the corticofugal fibers) which
increase alertness and decrease the desire to sleep ( during emotions and
voluntary movements)
3. Certain drugs called the analeptic drugs e.g. catecholamine’s,
amphetamine.
B. Factors that decrease the ARAS activity
1. Reduction of signals from either the sensory pathways or the cerebral
cortex
2. Stimulation of the sleep centers.
3. Extensive damage of the ARAS (e.g. by tumors’)
4. General anesthetic drugs: these drugs lead to unconsciousness through
depressing the ARAS activity. They inhibit the synaptic transmission
between its neurons (by producing a state of hyper polarization in these
neurons.
166
Physiology
Dr. Basim Mohamad Alwan Lecture 17
THE LIMB IC SYSTEM
COMPONENTS OF THE LIMBIC SYSTEM
The limbic system is the area of the brain that regulates emotion and
memory. This system consists of a group of structures that make the border
between the neocortex and the brainstem (limbus = border). It has two
components:
1. The limbic lobe (archicortex and paleocortex) (fig. 17-1). It is the oldest part
of the cortex. It includes the subcallosal gyrus, the cingulate gyrus, the
retrosplenial cortex, the hippocampus, the parahippocarnpal gyrus and the uncus.
It also includes the piriform cortex and the entorhinal cortex which are connected
to the olfactory bulb and tubercle.
2. A group of deep structures intimately related to the limbic lobe.
Figure 17-1: A view of the medial surface of the right cerebral hemisphere showing the
components of the limbic system
They include the hypothalamus, the amygdale, and the anterior nuclei of the
thalamus, the septal nuclei and the upper part of the midbrain (the limbic
midbrain area - LMA) (fig. 17-2).

167
Figure 17 – 2: The limbic system.
M.Str and, LStr; medial and lateral olfactory striae; Str med; stria medullaris; Tub; olfactory
tubercle; DB; diagonal band of Broca; Sep; septum; AT; anterior nucleus of the thalamu; M;
mamillary body; H; Habenula; P; interpeduncular nucleus; MFB medial forebrain bundle;
CONNECTIONS (fig.17-3)
Besides the extensive interconnections between different parts of the limbic system,
it is also connected to higher and lower centers.
The temporal cortex mediates information from the visual, auditory, and somatosensory
cortices to the amygdala and the hippocampus. The orbitofrontal cortex is connected to
the hypothalamus. It is the only neocortical region with direct connection to the
hypothalamus and the most important neocortical control element on the limbic
functions.
The medial forebrain bundle connects the thalamus and hypothalamus with the
orbitofrontal cortex upwards and with the limbic midbrain area (LMA) downwards.
PAPEZ CIRCUIT:
and is chiefly involved in the
control of
The Papez circuit plays a role in storing memory. Described by
1937, Papez discovered the circuit after injecting
and monitoring its progression through the brain.
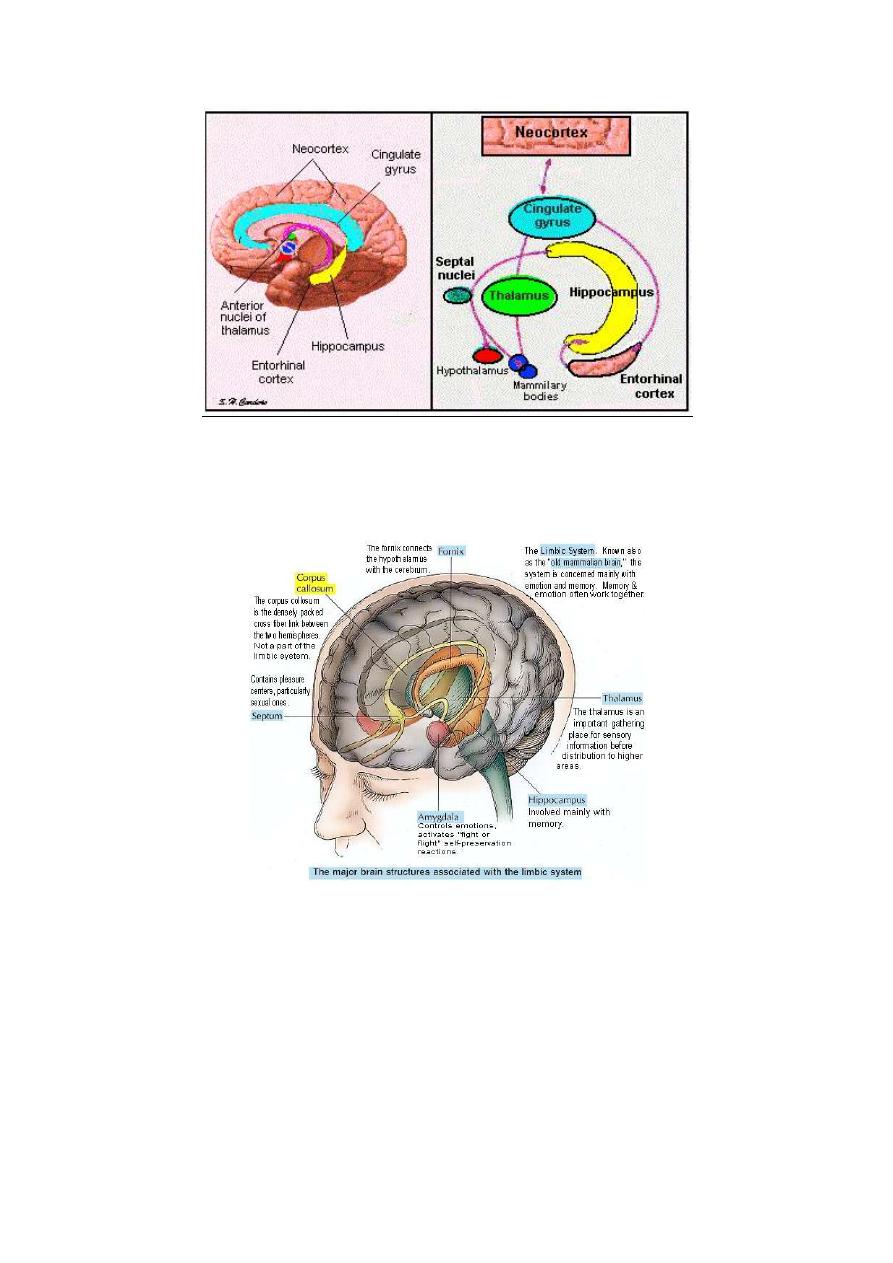
168
Fig. 17-3 Papez circuit
Fig. 17-4 the brain structures associated with the limbic system
CHARACTERISTIC FEATURES OF THE LIMBIC SYSTEM
• It contains a large number of excitatory neuronal circuits that act as reverberating
circuits.
• Impulses that excite the system produce a prolonged after discharge in the system
i.e. the response outlast the stimulus.

169
• Its functions are closely associated with hypothalamic functions.
THE REWARD AND PUNISHMENT SYSTEMS
The limbic system contains reward and punishment systems. The reward system
consists of a major reward center in the lateral and ventromedial nuclei of the
hypothalamus, and less potent centers in the amygdala, the thalamus and the
tegmentum of the limbic midbrain area. Stimulation of the system gives a sense
of reward; i.e. relaxation, pleasure and satisfaction. The subject is motivated to
approach or repeat the rewarding experience (the approach reaction).
The punishment system consists of a major punishment center in the periventricular
nucleus of the hypothalamus which extends up to the thalamus and down to the
periaquiductal gray matter of the limbic midbrain area, and other less potent centers
in the amygdala and the hippocampus. Stimulation of the punishment system gives a
sense of punishment i.e. displeasure, fear, and terror. The subject is motivated to avert
the punishing experience (the aversion reaction).
Dopamine is the main chemical transmitter in the reward system. Cocaine produces a
sense of well-being and pleasure by increasing the release of dopamine from the
dopaminergic nerve endings in this system.
The reward and punishment systems are very important for motivation, emotions,
memory, and learning. Any experience that stimulates either of these systems can be
easily admitted into long-term memory. Without stimulating the reward or
punishment systems, the experience has to be rehearsed or practiced repeatedly before
it could be stored in the long-term memory.
FUNCTIONS OF THE LIMBIC SYSTEM
1. OLFACTION
Olfaction is the oldest function of the limbic system. The limbic system is
concerned with perception and discrimination of different odors. It stores olfactory
memories and controls the emotional responses to olfactory stimuli.

170
2. MOTIVATION
Motivation is the feeling which activates a certain behavior to achieve a certain goal.
Motivation is controlled by the limbic association area. The reward and punishment
systems help the limbic association area to decide whether to approach or avert a
certain experience.
3. EMOTIONS
Emotions are states of strong feelings, associated with autonomic, and endocrinal
changes, and a strong affect. An emotional state starts by stimulation of the reward
or the punishment systems. This activates the excitatory reverberating circuits of
the limbic system which set other structures into action:
1. The thalamus which activates the cerebral cortex and increases
the level of alertness.
2. The hypothalamus which starts autonomic and endocrinal reactions; e.g.
tachycardia, hypertension and adrenaline secretion.
3 The amygdaloid nuclei which adjust the hypothalamic sensitivity
4. The orbitofrontal cortex which decides the behavioral motor response to the
incoming signal.
5. The hippocampus which sorts out the input signals and decides whether they
will be stored in memory or ignored and discarded.
Emotions outlast the provoking stimuli because they depend on the activity of the
excitatory reverberating circuits of the limbic system which have a long
afterdischarge. Emotions also cannot be turned on or off voluntarily because the motor
cortex has no efferent connections that can interfere with these circuits.
Stimulation of the medial, forebrain bundle or the septal nuclei produces a sense
of joy and happiness.
4. MEMORY
The limbic system plays an important role in sorting out the sensory signals into
significant ones which are stored in memory and insignificant ones which are

171
ignored. It is also important for encoding and consolidation of memory.
5. LEARNING
The hippocampus and amygdala are quite important for operant conditioning and
intelligence.
6. CONTROL OF FEEDING BEHAVIOR
The amygdaloid nucleus "sorts out" food into edible and inedible material on the basis
of past experience. Lesions in the amygdaloid nuclei lead to hyperphagia. However,
unlike animals with lesions in the hypothalamic satiety center, animals with
amygdaloid lesions try to eat any available object (omniphagia) before they discover
whether it is edible or not (oral exploration).
7. CONTROL OF SEXUAL BEHAVIOR
Sexual behavior in man is largely controlled by the orbitofrontal cortex. But the
instinctual desires and innate reactions that lead to mating and pregnancy are
functions of the limbic system and hypothalamus .
Lesions in the piriform cortex in the periamygdaloid area produce hypersexuality in
male animals. The sexual desire is intensified; the animals copulate with adult and
non-adult females of the same and of other species. They even mount male animals
and inanimate objects. The same lesions in female animals do not affect their
sexual behavior.
8. CONTROL OF MATERNAL BEHAVIOR
Nursing and care of young babies is an instinctual behavior in the female. The
instinctual maternal feelings and behavior are facilitated by the hormone prolactin.
They are markedly depressed by lesion in the cingulate and retrosplenial gyri.
9. CONTROL OF AUTONOMIC FUNCTIONS
Simulation of limbic structures produces autonomic effects; e.g. changes in heart

172
rate and arterial blood pressure. The response produced from wide areas in the
limbic system. It is part of response to emotions.
10. CONTROL OF RELEASE OF SOME HORMONES
Catecholamines and ACTH are released in response to stimulation different areas of
the limbic system. It is part of the response to emotions (the stress or alarm
response).
173
PHYSIOLOGY
Dr. Basim Mohamad Alwan Lecture 18
SLEEP
Sleep is a state of unconsciousness from which the person can be aroused by
sensory stimuli. A normal adult person sleeps 7-8 hr/day.
A newly-born infant sleeps much longer time (16-18 hi/day), whilst an old person
sleeps less (5-6 hr/day).
Figure 18 - 1: sleep cycle
WAKING / SLEEPING RHYTHM
Waking and sleeping periods follow each other in a circadian rhythm (i.e. 24 hr.
rhythm) which is synchronized with the daily light-dark cycle. This synchronization is
the function of the suprachiasmatic nucleus of the hypothalamus which receives
collateral from the visual pathway Physical and psychological factors affect the
onset and duration of sleep. E.g. cold and fear prevent the onset of sleep, whilst
fatigue and boredom facilitate its onset. In days preceding examinations, students
may use sleep as a legitimate escape way to avert studying.

174
TYPES OF SLEEP
There are two types of sleep which follow each other in a cyclic manner during
the sleep period; these types are:
1. NON-REM SLEEP
Non-REM sleep is also called slow wave sleep, or NREM sleep. It is a quiet sleep
during which there is no REM (Rapid Eye Movement). There are no dreams, but
other signs of mental activity might show up; as sleep-talking or sleep walking
(somnambulism) and night terror of children. Muscles are relaxed.
There are four stages of the non-REM sleep which can be identified by the EEG
recording as follows.
(a) STAGE I (very light steep)
This stage is characterized by slowing of the EEG rhythm and the appearance of
small theta waves. If a person is awakened during this stage, he asserts that he
was not asleep, but just closing his eyes. This stage occurs only at the onset and at
the waking up from sleep.
(b) STAGE II (light sleep)
This stage is characterized by the appearance in the EEG of sleep spindles on a
background of theta waves. Sleep spindles are bursts of large waves with a
frequency of 14-15 Hz which last for 1-2 seconds.
(c) STAGE III (intermediate sleep)
This stage is characterized by the appearance in the EEG of rapid delta waves
(frequency 3-3.5 Hz). Breathing is slow and even, pulse rate is about 65 beats
/min, temperature and arterial blood pressure continue to decline.
(d) STAGE IV (deep sleep)
This stage is characterized by the appearance in the EEG of delta-max waves
(delta waves with maximum slowing; frequency 0.5 - 1.2Hz). This stage occurs
only in the early sleep cycles. There is maximum slowing of breathing and heart
rate (about 60 beats/min). Cheyne-Stoke breathing may occur.

175
2. REM SLEEP
During REM {Rapid Eye Movement) sleep, there is rapid roving movement of the
eye. Dreams occur during this type of sleep. The EEG shows the desynchronized
β-rhythm. Although the EEG pattern is indicative of an activated brain, the
awakening threshold is as high as in deep sleep. That is why REM sleep is also called
"paradoxical sleep". There is marked decrease in muscle tone except for occasional
twitches of the facial and finger muscles. During REM sleep, dreams with sexual
elements are associated with erection of the penis and possible ejaculation.
SLEEP CYCLES
Normal sleep starts by non-REM sleep for about 80 min (70- 100) followed by
REM sleep for about 20 min (10-40). The cycle is then repeated but with shorter
NREM and longer REM periods. Four to five sleep cycles occur in a normal one
night sleep. In the later cycles, sleep becomes lighter with no NREM stage IV
(deep sleep) phases. The duration of deep sleep periods shortens with the advance
of age.
MECHANISM OF SLEEP
There are three theories to explain how sleep is induced. All of them are valid and
operating for induction of sleep and controlling the waking/sleeping rhythm.
1. THE METABOLIC THEORY
During wakefulness, brain cells produce a sleep-inducing factor (factor-S) which
accumulates in the CSF. When it reaches a certain level it induces NREM sleep.
This factor (a glucopeptide) was isolated from the CSF and urine of persons after
a period of sleep deprivation. When injected in humans of animals, factor-S
induces NREM sleep. A REM sleep factor also exists. Serotonin is considered as
a "sleep hormone" because it stimulates the production of these sleep-inducing
substances. A deep sleep-inducing peptide (DSIP) was isolated from the CSF and

176
urine of subjects during deep sleep.
Table 18 -1: A comparison between NJREM and REM sleep.
Criteria
NREM sleep
REM sleep
1. Incidence
Before REM sleep
After NREM sleep
2. Duration (per cycle)
Longer (80 rnin)
Shorter (20 min)
3, Depth of sleep
Variable
Very light
4. Threshold of
awakening stimulus
Variable
High
5. Rapid eye movement
Absent
Present
6. Dreams
Absent
Present
7. Muscle tone
Low
Very Low
8. Sleep talking & sleep
walking
May occur
Do not occur
9. Penile erection
Absent
Present
10. EEG
8- & δ-waves
β-waves
Nucleus ceruleus
11, Controlling center
Raphe nuclei
The concentration of factor-S in the brain declines steadily during sleep leading
finally to termination of sleep and start of wakefulness. According to the
metabolic theory of sleep, the important factor in determining the
waking/sleeping rhythm.
2. THE PASSIVE THEORY (DEAFFERENTATION THEORY)
The ascending reticular activating system (ARAS) sends facilitatory signals to the
cerebral cortex to increase its excitability and maintain the wakeful, alert state.
According to the passive theory, sleep is induced when the facilitatory signals from
the ARAS to the cortex are withdrawn. This occurs when the activity of the
ARAS is depressed either by fatigue or by lack of sensory input signals or
corticofugal signals.
This theory explains how sleep is rapidly induced by physical and mental

177
relaxation in a comfortable bed in a quiet, dark room at comfortable temperature.
Under these conditions, all sensory signals are reduced to minimal (functional
sensory deafferentation) and the ARAS activity is markedly reduced. Mental
relaxation eliminates any excitatory corticofugal signals and helps the onset of
sleep.
After a long period of wakefulness ARAS activity is depressed by fatigue. This
reduces the excitability of the cortical neurons leading to sleep.
3. THE ACTIVE THEORY (SLEEP CENTERS)
According to this theory there are specific centers which induce NREM sleep,
others which induce REM sleep. There is awaking/sleeping oscillator which
regulates the activity of these sleep centers.
Sleep is induced by induction of NREM sleep, REM sleep follows automatically.
(a) NREM SLEEP CENTER
The raphe magnus nuclei of the upper medulla and lower pons are considered as a
NREM sleeping center. Their stimulation induces NREM sleep. Their damage leads
to prolonged insomnia.
Inhibitory fibers from the raphe nuclei project to the ARAS and the cerebral
cortex. These fibers are serotonergic so drugs that block the synthesis of
serotonin, as chlorphenylalanin, produce prolonged insomnia. This insomnia can
be treated by 5-hydroxtrytophan. This is a precursor of serotonin, but unlike
serotonin, it can easily cross the blood-brain barrier.
(b) REM SLEEP CENTER
The nucleus ceruleus of the pons is considered as a REM sleep center. Its
stimulation converts NREM to REM sleep. It stimulates the cerebral cortex and
inhibits the raphe nuclei. The EEG shows the waking desynchronized β-rhythm
although the subject is asleep. It inhibits the facilitatory reticular formation
leading to marked decrease in the skeletal muscle tone. A lesion in the nucleus
ceruleus abolishes REM sleep, but NREM sleep can still occur.

178
(c) WAKING/SLEEPING OSCILLATOR CENTER
The suprachiasmatic nucleus of the anterior hypothalamus is responsible for
synchronizing the waking/sleeping rhythm with the 24-hr light/dark cycle. It is
considered as the waking/sleeping oscillator center. The suprachiasmatic nucleus acts
by stimulating the raphe nuclei which in turn induce sleep. Damage of this nucleus
leads to intense wakefulness. This eventually leads to severe exhaustion which could
be fatal.
JET LAG
Jet lag is the delay of synchronization of sleep and other biological functions of
the body to a shift in the light-dark cycle.
Flying by modem fast jet planes causes shift of the light dark cycles; the day of
the flight is shortened by flying eastwards and is lengthened by flying westwards.
The circadian rhythm of sleep and other biological functions take one day per
time zone (one-hour shift) for readjustment to synchronize with the new
light-dark cycles. Readjustment is easier after flight to the west than to the east.
PHYSIOLOGICAL CHANGES DURING SLEEP
NERVOUS SYSTEM
Voluntary activity and sensory perception are abolished. The brain is still
receptive and shows evoked potentials in response to sensory stimuli as clicking
sounds although these sounds may fail to arouse the sleeper. Protective spinal
withdrawal reflexes are depressed but can still be elicited. Sleeping synchronizes
various nervous centers together. This synchronization is disturbed during the
long periods of wakefulness which leads to increase in the rate of error in judgment
and behavior. The synchronization of nervous centers is the main function of
sleep; i.e. putting all the nervous centers back to the zero line.
CARDIOVASCULAR SYSTEM
There is slowing of the heart down to 60 beats/min and drop in the arterial blood

179
pressure. Also, there is general slowing of the circulation with prolongation of the
circulation time.
RESPIRATORY SYSTEM
There is decrease in the rate and depth of breathing which decreases pulmonary
ventilation, Cheyne-Stoke's breathing may occur due to slow circulation and
prolonged lung-to-brain circulation time. Airway resistance increases because of
the increased vagal tone.
BLOOD
PCO
2
rises up with a tendency to acidosis due to decreased pulmonary ventilation.
The hematocrit value increases because of the chloride shift.
DIGESTIVE SYSTEM
There is enhanced secretory and motor activity. Splanchnic blood flow increases
and absorption is enhanced.
METABOLISM
Metabolic rate decreases by 10-30 %. Body temperature drops by 0.5- 1°C. The
circadian
metabolic
and
temperature cycles
are synchronized
with
waking/sleeping cycles, but are not dependent on sleep; i.e. they occur even
without sleep.
ENDOCRINE GLANDS
Secretion of most hormones show a variable degree of circadian rhythmicity
which is synchronized with the waking/sleeping rhythm but again it is mostly
independent of sleep. ACTH and Cortisol secretion decreases to a minimal level
at midnight and reaches a maximum level at about 8 AM.
Secretion of growth hormone, however, is directly related to sleep. It is stimulated
by NREM sleep and inhibited by REM sleep.
SKELETAL MUSCLE TONE
Skeletal muscle tone decreases during sleep, it decreases during REM more than
during NREM sleep. The decrease in muscle tone occurs by inhibition of the
facilitatory reticular formation.

180
KIDNEYS
The rate of urine formation increases due to the decrease in sympathetic tone and
the increase in the renal blood flow.
PUPILS
Pupils are constricted despite lid closure. The oculomotor nucleus is released from
cortical inhibition. The parasympathetic tone is higher
during sleep.
SLEEP DISORDERS
INSOMNIA
Insomnia means lack of sleep despite convenient surroundings. A normal adult
can manage with 5.5 hr of sleep/day for long periods without ill effects. Insomnia
may he caused by psychological factors as anxiety, fear, or depression; or the
intake of analeptic substances as tea, coffee, or cola; or lesions in the sleep centers
in the brainstem or the suprachiasmatic nucleus of the hypothalamus, or an
irritative lesion in the dorsal hypothalamus.
Prolonged insomnia leads to bizarre behavior, temporary neurosis and psychosis.
Mental brightness and memory fail with lack of attention. After 4-5 days of sleep
deprivation, hallucinogenic substance similar to LSD (lysergic acid diethylamide)
appears in blood leading to hallucinations.
SNORING
It is the production of sound by a sleeping person during breathing. The person is
usually in the supine position with the mouth open. The tongue sinks back into the
throat. Snoring is occasionally followed by sleep apnea which could be fatal.
SLEEPTALKING
This is not an abnormality. It is just a sign of mental activity which occurs during
NREM sleep.
SLEEPWALKING (Somnambulism)
This is neither pathological nor harmful symptom. It happens to people at any age

181
but is most common in children and young adults.
The eyes of the sleepwalker are open, gazing straight ahead to nothing. He avoids
obstacles in his way but pays no attention to the surroundings. Sleepwalking
occurs during deep NREM sleep and is not accompanied by dreaming. It is a
special form of walking guided by sensory stimuli but without consciousness.
BED WETTING (Enuresis)
This is a, reflex micturition that occurs during sleep without any control from the
higher centers. It occurs during deep NREM sleep with no dreams.
Psychological factors may contribute to this condition.
NIGHT TERROR OF CHILDREN (Pavor nocturnus)
In this condition, the child sits up shortly after sleep onset, screaming and
appearing to stare at someone or something with wide open eyes. The face is pale
and covered with sweat, and breathing is difficult. After a short time, the child
wakes up, recognizing his surroundings and, if reassured, goes back to sleep. This
condition occurs during deep NREM sleep. The child usually cannot recall any
dreams.

182
PHYSIOLOGY
Dr. Basim Mohamad Alwan Lecture 19
LEARNING
Learning is the ability of the brain to use previous experience to modify
the inborn reactions or create new ones. There are two types of
learning; non-associative and associative.
1. NON-ASSOCIATIVE LEARNING
In this type of learning, the subject learns whether to ignore or to respond to a
certain stimulus. This occurs through two mechanisms; habituation and
sensitization.
(a) HABITUATION
This is the gradual decrease in response when the stimulus is frequently repeated.
For example, a loud and unexpected sound produces several reflexes, i.e. looking
towards the sound source, change in heart rate and blood pressure. If the sound
turns out to be insignificant to the subject, subsequent repetition of the sound
produces little or no response at all.
Habituation is stimulus specific. If one is habituated to traffic noise, he will be
able to sleep in this noise. Then, on a background of traffic noise, either an
unusual sound or unusual silence will wake him up. Also, a mother who sleeps
through many kinds of noise, wakes up promptly when her baby cries.
Habituation is the simplest and most widespread form of learning. Through
habituation, one learns to ignore a huge number of insignificant stimuli.
(b) SENSITIZATION
This is the potentiation of a response to a certain stimulus by coupling it with
another intense or noxious stimulus. For example, one normally ignores stray

183
dogs (by habituation), but if one is bitten by a stray dog, he becomes very attentive
to avert dogs for a long time (the aversion reaction).
Sensitization is stimulus specific. A person who was bitten by a dog will not be
afraid of a caw.
Habituation and sensitization are simple but very important learning processes.
Through them, one learns to direct his attention to significant stimuli and ignore the
much more numerous insignificant ones.
2. ASSOCIATIVE LEARNING
In this type of learning, the subject learns the correlation between one stimulus
and another. Associative learning occurs by conditioning of body reactions.
There are two types of conditioning; classical and operant conditioning.
(a) CLASSSCAL CONDITIONING
Classical conditioned reflexes are reflexes in which a nonspecific stimulus is
made to produce the same response as the specific stimulus of a certain reflex.
They are produced by applying the nonspecific stimulus (conditioned stimulus -
CS) before the specific stimulus (unconditioned stimulus - US) for several times.
When conditioning is established, the application of the CS produces the same
response as the US.
For example, salivation is a normal response to food intake. If a person gets used
to taking tasty meals in a certain restaurant, when conditioning is established, the
entry into this restaurant would reflexly stimulate salivary secretion even without
taking food. In this case, the entry into the place which is the conditioned stimulus
(CS) becomes linked to food intake which is the unconditioned stimulus (US).
To establish a conditioned reflex, the CS must be applied before the US not after it.
The two stimuli must not be separated by a long interval or by any distracting
stimulus. The pairing of the CS and US must occur for several times until new
facilitated pathways for the new conditioned reflex are formed.
The center of classical conditioning is the cerebral cortex. Most of the classical

184
conditioned reflexes are integrated in the orbitofrontal cortex.
(b) OPERANT CONDITIONING
Operant conditional reflexes are "reflexes in which the subject learns to take an
action in response to a stimulus to get a reward or avert a punishment"; i.e. the
subject "operates" on his surroundings. For example, a car driver would slow
down and stop the car on seeing the red traffic light, and drives on when seeing the
green light.
Operant conditioning cannot be established in the absence of the cerebral cortex,
especially the orbitofrontal cortex. The hippocampus and the amygdala are
important in linking the stimulus to the operation.
INTELLIGENCE
Intelligence is the efficient employment of past and present experience to get the
best solution for a problem. It is a mental faculty which requires three brain
capabilities:
1. Rapid grasping of significant signals, their coding and storage in the
appropriate memory stores.
2. Rapid retrieval of the appropriate, sufficient information related to current
experience.
3. Efficient linking of information from past experience to information from the
current one to reach accurate evaluation of the situation and formulating or
creating ideas of the best way to handle it.
Although man-made computers grasp, store, retrieve and link data fed to them,
they differ from the brain in that they cannot create new ideas although they can
formulate solutions to problems based on the stored data.
Intelligence is a combined function of the cortical association areas together with
the limbic system. It is the basis for professional efficiency particularly in
intellectual professions as scientists and physicians. High intelligence requires
rich, well coded memory stores.

185
Intelligence is not inherited. It is an inborn brain capability which is influenced by
social and environmental conditions. It can be improved by appropriate training.
Trials to improve intelligence by drugs have all failed.
The intelligence quotient (IQ) is a number indicating the ratio of a person's
intelligence relative to the average.

186
PHYSIOLOGY
Dr. Basim Mohamad Awan Lecture 20
MEMORY
Memory is the ability of the brain to store information and retrieve it at a later
time. The storage capacity of the human brain is limited. So, the information that
flow into the brain is classified. The most important ones (less than 1%) are
selected and stored, but all the rest are neglected and forgotten.
STORAGE CAPACITY OF THE BRAIN
The information unit is the "bit", A bit is the simplest form of sensory experience;
i.e. a letter, a line, a color, a tone, a smell...etc. The capacity of all the sensory system
to send information to the brain is less than 50 bits/s. E.g. during quiet reading, the
rate of information flow to the brain is 40 bits/s, during mental calculation it is 12
bits/s and during counting it is 3 bits/s. An average rate of information flow is about
20 bits/s. For learning a language, 40-50 million bits should be stored in memory.
Ten neurons are required to store one bit of information. The total storage capacity
of the human brain is about 3x 10
8
bits.
TYPES OF MEMORY
There are four different types of memory:
[I]
SENSORY MEMORY (Immediate memory).
[II] PRIMARY MEMORY (Short-term memory)
[III] SECONDARY MEMORY (Long-term memory)
[IV] TERTIARY MEMORY (permanent memory)
[I] SENSORY MEMORY (Immediate memory)
Any sensory signal is automatically stored in the sensory memory for few seconds.
This memory can accommodate 15-20 bits. Forgetting starts spontaneously

187
immediately after the information is acquired. The spontaneous gradual decline in
the amount of stored information is called "fading" of information. The
spontaneous disappearance of information from the memory is called
"extinction" of information. The information in sensory memory can be
transferred to the secondary memory.
[II] PRIMARY MEMORY (Short-term memory)
This is a memory that lasts for few minutes to few hours. The information enters
this memory by verbalization; i.e. through spoken or written words. The primary
memory is not found in young infants or animals as they would not understand the
meaning of spoken or written words.
The capacity of primary memory is small. Bits of information are introduced into
it in chronological order, one after the other. The new information replaces the old
ones. The latter are then forgotten.
The access to the primary memory stores is rapid; one can retrieve the information
rapidly.
[III] SECONDARY MEMORY (Long-term memory)
This is a memory that lasts for several hours up to several years. The information
is introduced into this memory from the sensory and primary memories by two
mechanisms:
L Stimulation of the reward or punishment systems.
2. Repeated practice or rehearsal of the experience.
The capacity of the secondary memory is large. The information is stored
according to its significance. The bits of information of related significance are
stored together. The access to the secondary memory stores is slow; it takes some
time to remember the wanted information.
Forgetting of information in the secondary memory occurs when a new
information conflicts with an old stored one. In this case, one information cancels
the other either by repulsion or by replacement If the 'old information repels the
new one; the process is called "proactive inhibition". If the new information

188
replaces the old one, the process is called "retroactive inhibition".
In this way. the more we have learned, the greater the liability to forget the new
information by proactive inhibition. The old information inhibits the acquisition
of new ones. Therefore, the blame for most of our forgetting must be placed on
what we have learned before.
[IV] TERTIARY MEMORY (Permanent memory)
This is the permanent memory. The information stored in this memory are never
forgotten; e.g. one's name or the ability to read and write. Information in the
tertiary memory comes from the secondary memory by years of practice, which
strongly consolidates the memory. The stored information in the tertiary memory
remains available for retrieval even if information in other memories are erased
by brain injury or disease. This is because information in the tertiary memory
occupy large areas of the brain and more than one "copy" of the information are
stored in different regions of the brain.
The access to the ternary memory is very rapid, e.g. one immediately remembers
his name if he is asked about it.
MECHANISMS OF MEMORY
[I] SENSORY MEMORY
Sensory memory is made by prolonged after discharge in the neuronal circuits.
Three mechanisms are involved:
(a) Activation of reverberating circuits leading to repeated reactivation of
neurons.
(b) Short term synaptic potentiation by multiple successive stimulation of input
neurons.
(
C
)
Synaptic sensitization by coupling the sensory experience with intense or
noxious stimulus.

189
[II] PRIMARY MEMORY
The primary memory is made by the formation of temporary memory traces. A
memory trace is a newly developed pathway for signal transmission resulting
from facilitation of new synapses. This leads to the creation of new circuits in the
brain that keeps the memory of the experience. Activation of these circuits brings
the memory up to one's mind. There are two possible mechanisms for the
formation of the new memory traces:
(a) Long term potentiation of synapses. During verbalization the brain catches
the new interesting information and rehearses it several times.
(b) Changes in the physical properties of the postsynaptic membrane
leading to enhanced sensitivity to the chemical transmitter.
[III] SECONDARY MEMORY
Secondary memory is made by formation of "memory engrams". A memory
engram is a long-lasting memory trace formed by structural changes in
presynaptic terminals. These changes inc lude:
(a) Increase in the total area of the specific release sites in the cell
membrane. Transmitter vesicles get attached to these sites before they rupture
and release their transmitter into the synaptic cleft. The increase in the release
sites increases the amount of transmitter released at the presynaptic terminal, thus
facilitating synaptic transmission.
(b) Increase in the number of transmitter vesicles in the presynaptic
terminal.
(c) Increase in the number of presynaptic terminals by formation of new
terminals.
The memory engrams remain for a long time, up to several years. Formation of
memory engrams requires protein synthesis. Antibiotics which inhibit protein
biosynthesis interfere with the formation of memory engrams. They prevent the
consolidation of memory but do not affect the primary memory.
190
[IV] TERTIARY MEMORY
Tertiary memory is made by the formation of permanent memory traces
(permanent engrams). These permanent engrams are made by structural
changes in the neurons. The changes are similar to those in secondary memory.
Tertiary memory may be considered as an advanced stage of secondary memory.
Table 26-1 compares and contrasts the different types of memory
Table 19-1: Characteristics of different types of memory.
Sensory
Primary
Secondary
Tertiary
Capacity
Very small
Small
Very large
Very large
Duration
Few seconds
Minutes to hours
Hours to years
Permanent
Entry into storage Automatic during
perception
Verbalization
Practice or stim.
of reward or
punish, systems
Repeated practice
for long time
Speed of retrieval
Very rapid
Rapid
Slow
Very rapid
Type of stimulus
Sensory
Verbal
All forms
All forms
Forgetting
Spontan. fading &
extinction
New information
replaces the old
Pro.and retro.
active inhibition
No forgetting
CONSOLIDATION OF MEMORY
Consolidation of memory means the transfer of information from the Sensory and
primary short-term memories to the secondary long-term memory. This process takes
from 5 minutes for minimal consolidation to two hours for maximal consolidation.
Consolidation of memory could be interrupted by deep anesthesia, brain
concussion or electroconvulsive therapy (ECT). Accordingly, if a sensory
impression is made, then followed within 5 minutes by brain concussion or ECT or
anesthesia, the experience gets extinct. This explains why patients who had brain
concussion in accidents cannot remember what happened at the time of the

191
accident. If the concussion occurs 2 hours after" the sensory experience, the
information remains unaffected. If it occurs within two hours, the memory is
affected proportionately.
Consolidation occurs to the information which attracts the attention of the mind.
The brain automatically rehearses this information.
A wide awake person consolidates memories far better than a person with mental
fatigue. This is because the wide awake brain is more attracted by the new
information and is capable of making rapid and more frequent rehearsals that
would consolidate memory more effectively.
Normal quiet sleep consolidates the memory of information received before the
onset of sleep.
ENCODING OF MEMORY
Encoding of memory is the classification, then placing each memory item with other
related items in the proper memory store; i.e. memory archiving. It is part of the
consolidation process. The hippocampus plays a central role in this function. All bits
of information go first to the hippocampus where they are sorted out as significant or
insignificant. If the information is classified as significant, single are sent to the
mamillary bodies of the hypothalamus. From the hypothalamus signals proceed on
to the orbitofrontal cortex, then to the basal forebrain (fig. 19-1). From the basal
forebrain (nucleus basalis of Meynert) there are diffuse cholinergic projections to
the memory stores which are found in all parts of the neocortex, the amygdala and
the hippocampus.
The amygdala associates the memories formed through different senses, then
through its connections with the hypothalamus (the amygdalohypothalamic
pathways) it is responsible for the emotional and autonomic responses to
memories.
192
Figure 19-1: brain areas concerned with encoding of long term memory.
Long-term memories are stored in the form of engrams in different regions of the
brain. To retrieve a certain memory item, one should get an access to the specific
engrams of this item and activates them. This can be done by different
associations. For example the word "Egypt" could be accessed and activated by
other words as "the pyramids", "the sphinx", "the Nile" or hearing part of the
opera "Aida" or seeing a photo of a pharaoh with his characteristic features and
head dress. Each of these methods of access is considered as a specific "key"
which "unlocks " and activates the specific engrams of the stored item.
FACIAL RECOGNITION AREA
The impression effaces is stored in the "facial recognition area" in the
neocortex of the undersurface of the temporal and occipital lobes (fig. 19-2). In
right-handed persons and some of the left handed ones, the facial recognition area
is much better developed in the right (representational) hemisphere than in the left
(categorical) hemisphere.
Bilateral lesion in this area leads to "prosopagnosia" which is' inability to
recognize people by their faces. In this case, the patient can recognize persons by
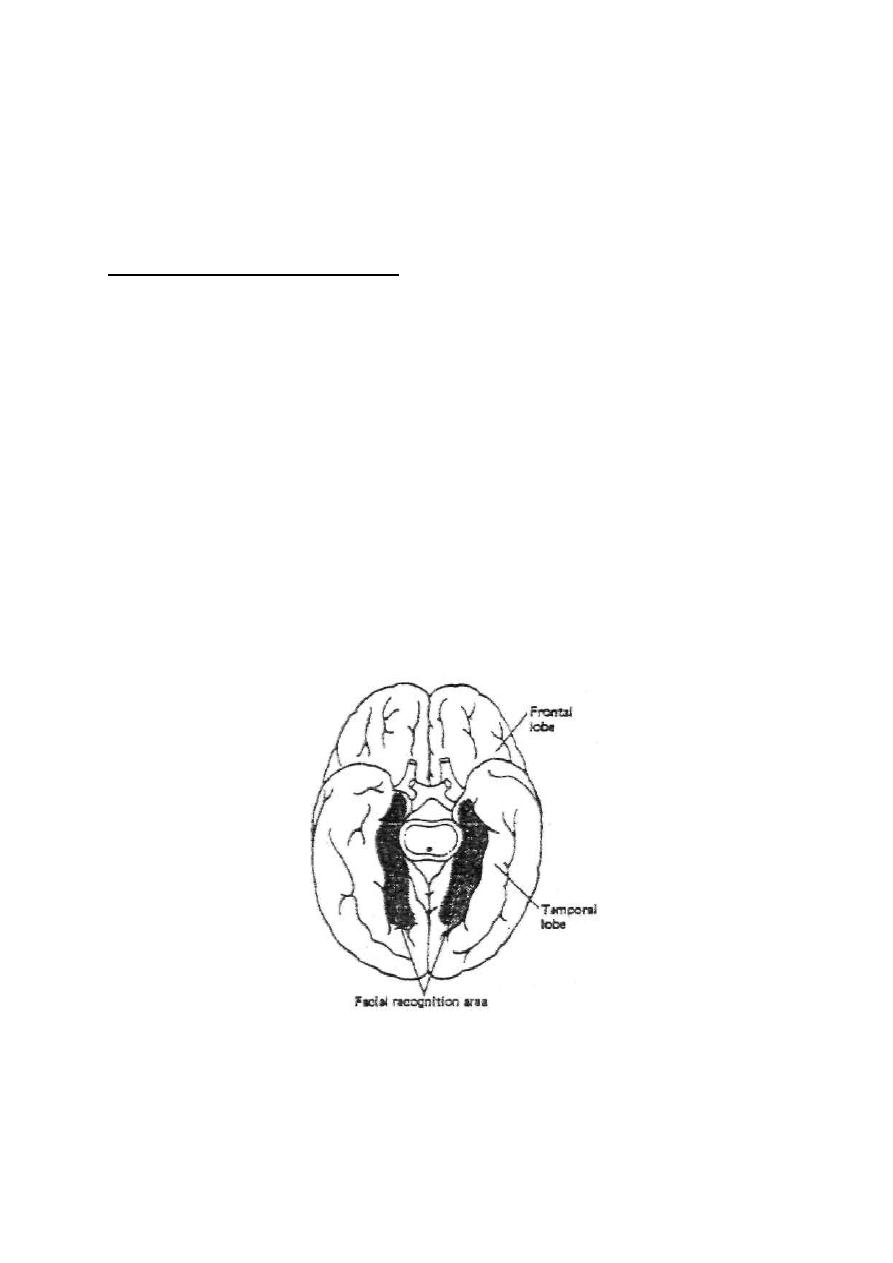
193
their voices. A prosopagnostic patient develops emotional and autonomic
reactions on seeing a familiar face, but he wouldn't identify the person until he
hears him speaking.
NAMING OF OBJECTS AREA
This is the function of an area in the most lateral portion of both the anterior
occipital and posterior temporal lobes. In this area, the names of different objects
are stored. Like the facial recognition area, it is better developed on the right side
than in the left side. Damage of this area leads to inability to name objects,
although the patient is able to know what the object is, its value or its use. Only
the name is lost from memory.
This is probably the brain area which gave Adam
the advantage over angels. According to the holy Qur'an, after the creation of
Adam, angels failed to name objects when Allah asked them to do, Adam by
the command of Allah, preceded and told the names of objects.
Figure 19-2: basal view of the human facial recognition area.

194
AMNESIA
Amnesia (Gr. = forgetfulness) means the inability to remember past experience.
There are several types of amnesia:
1. RETROGRADE AMNESIA
It is the inability to recall events that occurred shortly before the onset of brain
malfunction without affecting memories of the remote past. Retrograde amnesia
occurs with brain concussion (post-traumatic
amnesia), anesthesia, electric shock (therapeutic ECT or accidentally). In these
conditions, a transient brain malfunctioning erases the memories of events over a
long period before the onset of malfunctioning. During recovery, the length of the
period of amnesia shrinks progressively till it involves only several minutes.
2. ANTEROGRADE AMNESIA
It is the inability to form new memories. The memories consolidated before the
onset of amnesia are retained. The sensory and primary memories are functional
but cannot be consolidated.
Anterograde amnesia occurs with bilateral lesions in the hippocampus or other
structures involved in the encoding of memory.
3. PSYCHOGENIC OR HYSTERICAL AMNESIA
This is a rare condition characterized by sudden loss of memory for usually all
information in the secondary and tertiary memories. This usually follows a severe
psychological trauma or it may be an unconscious response to internal conflict or
an intolerable life situation. It is a purely functional disorder without any organic
disease. It can be differentiated from amnesia caused by damage or disease in
the brain tissue by three characteristics:
(I) All the personal data are forgotten, including the patient's own name.
(ii) The amnesia is not affected by key stimuli; seeing members of his

195
family, his friends or his personal effects.
(iii) The inability to remember past events persists although the patient
can make new memories.
Complete recovery of memory almost always occurs.
ALZHEIMER DISEASE AND SENILE DEMENTIA
Alzheimer disease is caused by degeneration of the cholinergic nerve fibers
which project from the nucleus basalis of Meynert to the neocortex, the
amygdala and the hippocampus. The disease is characterized by deterioration
of intellectual abilities as impairment of memory, lack of judgment and
inattentiveness. The disease may occur at any age. In old age, it is called senile
dementia. It is the commonest cause of dementia in old age (found in 10-15% of
the population above the age of 65). The anticholinesterase drug eserine
(physostigmine) produces some improvement but it does not stop the progress of
the disease.

196
PHYSIOLOGY
Dr. Basim Mohamad Alwan Lecture 21
SPEECH
Speech is the expression of ideas by spoken words. It is a sophisticated function
which is a characteristic of the human brain.
LATERALIZATION OF THE SPEECH FUNCTION IN THE BRAIN
About 95% of the human populations are right handed. In those individuals, and
in some left-handed ones, the centers of speech are found in the left cerebral
hemisphere. The hand skills area is much better developed in the left hemisphere
as well.
It is because of this functional lateralization that the left hemisphere in those
individuals was called the "dominant" hemisphere. In the rest of the population,
the right hemisphere was the "dominant" one. However, the other hemisphere
cannot be called "nondominant" because it is more superior or "dominant" in
other functions as recognition of faces, stereognosis, recognition of musical
themes and spatial recognition of the body, That is why when speaking of
dominance it is better to talk about the "speech-dominant" or the "categorical"
hemisphere. The other hemisphere is better called the "nonspeech-dominant"
or the "representational" hemisphere.
* Categorical: Decisive; it takes important decisions and issues orders for actions
by the body.
* Representational: In which high sensory functions are represented, but with much
less role in making decisions or issuing of orders.

197
HEARING AND SPEECH
The development of the faculty of speech depends first on the ability to hear
spoken words. A young baby starts first to learn the meaning of the heard words
then tries to imitate them by vocalization (uttering simple sounds) then by
verbalization (uttering words). So, speech is learned basically through hearing. If
a person is born deaf, he cannot develop the faculty of speech and is destined to
be dump.
Perception of spoken words is the function of the primary auditory area (area. 41)
in the temporal lobe at the floor of the lateral sulcus. Signals are then conveyed to
the adjacent auditory interpretative area (area 42). This area understands the
meaning of the heard words. It feeds the message of the heard words to the
angular gyrus (general interpretative area - area 39) which correlates them with
other related items stored in memory in the process of thinking.
READING AND SPEECH
Written words are perceived by the primary visual area (area 17) in the occipital
lobe. The signals are then conveyed to the visual interpretative area (area 18 - 19)
which understands the meaning of words. The message is then fed to the angular
gyrus which correlates them with other related items stored in memory in the
process of thinking.
WERNICKE'S AREA (The language and speech center)
Wernicke's area is found in the posterior part of the superior temporal gyrus (fig.
20-1). It receives input signals from the angulr gyrus (general interpretative area -
area 39). It is the memory store tor language. It decides what words are suitable
and in what sequence to express a certain idea.
Wernicke's area is very well developed in the categorical but not in the
representational hemisphere. It is connected with Broca's area (word formation
198
center) in the premotor cortex via the "arcuate fasciculus.
THE WORD-FORMATION CENTER (Broca's area, area 44)
This area is found in the premotor area. It stores the motor programs for
different words. It receives input signals from Wernicke's area through the
arcuate fasciculus. When it is activated it stimulates the motor cortex at a certain
pattern to produce words by coordinated contractions of the respiratory,
laryngeal, pharyngeal, lingual and labial muscles.
Figure 20-1: Wernicke's area, angular gyrus
,
the arcuate fasciculus, and
Broca's area.
THE WRITING CENTER (Earner's center)
This is part of the hand-skills area which stores the motor programs for writing of
words or drawing of figures by the muscles of the hand. It receives input signals
from Wernicke's area via the arcuate fasciculus.
THE MECHANISM OF SPEECH
Speech passes by four steps to occur :
1. Formation of thoughts and ideas that will be expressed in speech: This is
the function of the angular gyrus. The ideas are formed in response to signals
from the visual, auditory, and other sensory areas.
2. Choice of suitable sentences and phrases to express the ideas:

199
This is the function of Wernicke's area which receives input signals from the
angular gyrus and sends output signals to Broca's area.
3. Word formation: This is the function of Broca's area which receives input
signals from Wernicke's area and sends programmed impulses to the primary motor
cortex to move the muscles of speech in a specific sequence to produce different
words.
4, Verbalization: It is the coordinated contraction of muscles of speech in a
certain sequence to produce spoken words. This is the function of the motor
cortex, the motor nerves and the muscles of speech.
Ideas may be expressed in writing. In this case, Wernicke's area feeds the
signals to Exner's center to write the desired words.
SPEECH DEFECTS
Speech defects are divided into two main categories"
(I) Aphasia
(II) Dysarthria
(I) Aphasia
It is a speech defect in a patient who can see, hear and move the muscles of
speech.
TYPES OF APHASIA
1-Sensory (receptive) aphasia.
2. Motor aphasia.
3. Wernicke's aphasia.
4. Global aphasia.
1. SENSORY (RECEPTIVE) APHASIA

200
Sensory aphasia is found in two forms:
(a) Sensory auditory aphasia (word deafness): This is due to a lesion in the
auditory interpretative area. The patient can hear the spoken words, but is unable
to understand the message.
(b) Sensory visual aphasia (word blindness or dyslexia): This is due to a
lesion in the visual interpretative area. The patient can see the written words but
is unable to understand the message.
2. MOTOR (EXPRESSIVE) APHASIA
Motor aphasia is found in two forms:
(a) Vocal aphasia: This is due to a lesion in Broca's area. The
P
atient is
perfectly capable of deciding what he wishes to say and is capable of
vocalization, but he simply cannot make his vocal system emit words, but only
noises (nonfluent aphasia).
(b) Writing aphasia (agraphia):
This is one to a lesion in Exner's center. The
patient knows what he wants to writs or draw and he is capable of moving the hand
voluntarily, but he cannot make his hands write words or draw graphs to express
his thoughts.''
3. WERNICKE'S APHASIA
This aphasia is caused by lesions in Wernicke's area of the categorical
hemisphere. The patient comprehends ideas expressed by spoken or written
words. He formulates thoughts and ideas but doesn't know how to express them.
The process of verbalization is normal, so the patient can talk and sometimes he
talks too much (fluent aphasia). However, his speech is full of jargon
(speech
full of too much unnecessary words)
and neologism
(new words invented by
the patient)
that make little sense.
Very rarely, when the lesion is in the arcuate fasciculus, the Wernicke's, area is

201
disconnected from Broca's area. The patient knows what he wants to say, but
cannot produce the required speech. This condition is called "conduction
aphasia".
4. GLOBAL APHASIA
This aphasia is caused by extensive lesions in the categorical hemisphere
involving both frontal and temporal lobes. The aphasia is general; it involves the
receptive and expressive functions.
[II] DYSARTHRIA
Dysarthria means difficulty in producing clear, normal speech due to a defect in
the motor system of verbalization. There is either weakness, incoordination,
hyper or hypotonia, or paralysis of the muscles of speech.

202

203