
roots. After entering the cord, every sensory signal travels to two separate
The cord gray matter is the integrative area for the cord reflexes. Figure 54–1
Organization of the Spinal Cord
sic excitatory motor functions of the cord itself.
sensory input signals to the cord. Using this preparation, one can study the intrin-
reflexes become very excitable and, therefore, easy to activate by even the slightest
the spinal cord motor control circuits. The result is that the spinal cord motor
these nuclei to become tonically active, transmitting facilitatory signals to most of
centers of the brain to the pontile and vestibular muscle control nuclei. This allows
blocks normal inhibitory signals
encephalic level, which
study.
a few days to weeks in monkeys, most of the intrinsic spinal cord functions return
the level of transection is severely depressed. After a few hours in rats and cats or
still remains functional, and (2) the
which the spinal cord is transected, frequently in the neck so that most of the cord
been especially useful in studying spinal cord function: (1) the
Two types of experimental preparations have
circuits provide all but a small fraction of the direct control of the muscles.
ronal circuits of the spinal cord that are the objects of the commands. These
“command” signals generated in the brain. But it also requires the many neu-
tinuously and control equilibrium. All this is done through “analytical” and
change the movements from walking to jumping as needed, and to monitor con-
ments when they are required, to lean the body forward during acceleration, to
Let us not belittle the role of the brain, however, because the brain gives
signals to the spinal cord to set into motion the walking process.
Instead, the circuits for these movements are in the cord, and the brain simply
causes the specific to-and-fro movement of the legs that is required in walking.
To give an example, there is no neuronal circuit anywhere in the brain that
Without the special neuronal circuits of the cord, even the most complex motor
In this chapter, we discuss the control of muscle function by the spinal cord.
brum, where the most complicated muscle skills are
plicated responses, and finally extend to the cere-
reflexes, extend into the brain stem with more com-
We have already noted that sensory information is
Cord; the Cord Reflexes
C
H
A
P
T
E
R
5
4
673
Motor Functions of the Spinal
integrated at all levels of the nervous system and
causes appropriate motor responses that begin in
the spinal cord with relatively simple muscle
controlled.
control systems in the brain could not cause any purposeful muscle movement.
sends command
directions that control the sequential cord activities—to promote turning move-
Spinal Animal and the Decerebrate Animal.
spinal animal, in
decerebrate animal, in which the brain stem is
transected in the middle to lower part of the mesencephalon.
Immediately after preparation of a spinal animal, most spinal cord function below
to nearly normal and provide a suitable experimental preparation for research
In the decerebrate animal, the brain stem is transected at the middle to lower mes-
from the higher control
for Motor Functions
shows the typical organization of the cord gray matter in a single cord segment.
Sensory signals enter the cord almost entirely through the sensory (posterior)

where they are appropriately processed. Thus, in
signals are transmitted first through interneurons,
the anterior motor neurons. Instead, almost all these
types of circuits. In this chapter, we examine many
diverging, converging, repetitive-discharge,
interneuron pool of cells of the spinal cord, including
chapter.
in Figure 54–1. The interconnections among the
directly with the anterior motor neurons, as shown
with one another, and many of them also synapse
times per second. They have many interconnections
neous activity and capable of firing as rapidly as 1500
small and highly excitable, often exhibiting sponta-
as numerous as the anterior motor neurons. They are
shown in Figure 54–1. These cells are about 30 times
horns, and the intermediate areas between them, as
the cord gray matter—in the dorsal horns, the anterior
later in this chapter.
which helps control basic muscle “tone,” as discussed
muscle spindle,
shown in Figure 54–2. These
intrafusal fibers,
eter, which go to small, special skeletal muscle fibers
motor nerve fibers, averaging 5 micrometers in diam-
impulses through much smaller type A gamma (A
anterior horns. These gamma motor neurons transmit
muscle fibers, about one half as many much smaller
neurons, which excite contraction of the skeletal
Gamma Motor Neurons.
Transmission of
three to several hundred skeletal muscle fibers, which
innervate the large skeletal muscle fibers. Stimulation
averaging 14 micrometers in diameter; these fibers
) motor nerve fibers,
rise to large type A alpha (A
The alpha motor neurons give
Alpha Motor Neurons.
neurons.
of two types,
innervate the skeletal muscle fibers. The neurons are
They give rise to the nerve fibers that leave
neurons.
interneurons.
are of two types: (1)
discussed in Chapters 47 and 48, the other neurons
gray matter. Aside from the sensory relay neurons
bral cortex, as described in earlier chapters.
in the cord itself, to the brain stem, or even to the cere-
local effects. (2) Another branch transmits signals to
destinations: (1) One branch of the sensory nerve ter-
The Nervous System: C. Motor and Integrative Neurophysiology
674
Unit XI
minates almost immediately in the gray matter of the
cord and elicits local segmental cord reflexes and other
higher levels of the nervous system—to higher levels
Each segment of the spinal cord (at the level of
each spinal nerve) has several million neurons in its
anterior motor neurons and (2)
Anterior Motor Neurons.
Located in each segment of the
anterior horns of the cord gray matter are several
thousand neurons that are 50 to 100 per cent larger
than most of the others and are called anterior motor
the cord by way of the anterior roots and directly
alpha motor neurons and gamma motor
a
branch many times after they enter the muscle and
of a single alpha nerve fiber excites anywhere from
are collectively called the motor unit.
nerve impulses into skeletal muscles and their stimu-
lation of the muscle motor units are discussed in
Chapters 6 and 7.
Along with the alpha motor
gamma motor neurons are located in the spinal cord
g)
called
fibers constitute the middle of the
Interneurons.
Interneurons are present in all areas of
interneurons and anterior motor neurons are respon-
sible for most of the integrative functions of the spinal
cord that are discussed in the remainder of this
Essentially all the different types of neuronal cir-
cuits described in Chapter 46 are found in the
and other
applications of these different circuits in the perform-
ance of specific reflex acts by the spinal cord.
Only a few incoming sensory signals from the spinal
nerves or signals from the brain terminate directly on
Sensory root
Solitary cell
External basal cells
Corticospinal tract
Interneurons
Anterior motor
neurons
Motor root
cord.
with the interneurons and anterior motor neurons of the spinal
Connections of peripheral sensory fibers and corticospinal fibers
Figure 54–1
Ia
II
m
17
Motor
Sensory
Motor
Sheath
Primary
Alpha motor
ending
Gamma motor
ending
Fluid
cavity
ending
1 cm
Extrafusal
fibers
Secondary
ending
Intrafusal
fibers
14
m
m
m
5
m
m
g
g
8
m
m
a
5
m
m
tal muscle fibers. Note also both motor and sensory innervation of
Muscle spindle, showing its relation to the large extrafusal skele-
Figure 54–2
the muscle spindle.

type Ia fiber averaging 17 micrometers in diameter,
This nerve fiber is a
annulospiral ending.
of each intrafusal fiber, forming the so-called
large sensory nerve fiber encircles the central portion
In the center of the receptor area, a
secondary ending.
central receptor area of the muscle spindle. They are
Two types of sensory endings are found in this
of the spindle and therefore excites the receptor.
spindle’s intrafusal fibers stretches the midportion
change, contraction of the end portions of the
2. Even if the length of the entire muscle does not
the receptor.
midportion of the spindle and, therefore, excites
1. Lengthening the whole muscle stretches the
stretching of this midportion of the spindle. One can
fibers originate in this area. They are stimulated by
Figure 54–2 and in more detail in Figure 54–3, sensory
myosin and actin contractile elements. As shown in
this area, the intrafusal muscle fibers do not have
portion of the muscle spindle is its central portion. In
The receptor
Sensory Innervation of the Muscle Spindle.
muscle.
(type A alpha
gamma efferent fibers,
as described earlier. These gamma motor nerve fibers
that originate from small type A gamma motor
receptor, as described later. The end portions that
when the ends do. Instead, it functions as a sensory
ments.Therefore, this central portion does not contract
of these fibers—that is, the area midway between
muscle fiber. However, the central region of each
fibers.
54–2. Each spindle is 3 to 10 millimeters long. It is built
organization of the muscle spindle is shown in Figure
The
Structure and Motor Innervation of the Muscle Spindle.
Receptor Function of the
subconscious level. Even so, they transmit tremendous
muscle control. They operate almost completely at a
The signals from these two receptors are either
(see Figure 54–7), which are located in
length or rate of change of length, and (2)
(see Figure 54–2), which are distrib-
dantly with two special types of sensory receptors: (1)
tion, the muscles and their tendons are supplied abun-
length or tension changing? To provide this informa-
is its instantaneous tension, and how rapidly is its
instant. That is, what is the length of the muscle, what
information from each muscle to the spinal cord, indi-
Roles in Muscle Control
Tendon Organs—And Their
Muscle Sensory Receptors—
limbs.
later in this chapter, including reflexes that coordinate
transmit signals to many segments. These ascending and
transmit signals to only a segment or two, while others
both up and down the spinal cord; some of the branches
from the posterior cord roots, they bifurcate and branch
another. In addition, as the sensory fibers enter the cord
These fibers run from one segment of the cord to
propriospinal fibers.
Multisegmental Connections from One Spinal Cord Level
signals to spread laterally.
desired direction while suppressing the tendency for
sensory system uses the same principle—that is, to allow
focus, or sharpen, its signals in the same way that the
reason: The motor system uses this lateral inhibition to
This effect is important for the following major
adjacent motor neurons, an effect called
Thus, stimulation of each motor neuron tends to inhibit
mit inhibitory signals to the surrounding motor neurons.
cent Renshaw cells. These are
neuron, collateral branches from the axon pass to adja-
Renshaw cells.
motor neurons, are a large number of small neurons
horns of the spinal cord, in close association with the
Renshaw Cell Inhibitory System.
rons, where the signals from this tract are combined
Figure 54–1, the corticospinal tract from the brain is
Motor Functions of the Spinal Cord; the Cord Reflexes
Chapter 54
675
shown to terminate almost entirely on spinal interneu-
with signals from other spinal tracts or spinal nerves
before finally converging on the anterior motor
neurons to control muscle function.
Also located in the anterior
called
Almost immediately after the
anterior motor neuron axon leaves the body of the
inhibitory cells that trans-
lateral inhibi-
tion.
unabated transmission of the primary signal in the
to Other Levels—Propriospinal Fibers
More than half of all the nerve fibers that ascend and
descend in the spinal cord are
descending propriospinal fibers of the cord provide
pathways for the multisegmental reflexes described
simultaneous movements in the forelimbs and hind
Muscle Spindles and Golgi
Proper control of muscle function requires not only
excitation of the muscle by spinal cord anterior motor
neurons but also continuous feedback of sensory
cating the functional status of each muscle at each
muscle spindles
uted throughout the belly of the muscle and send
information to the nervous system about muscle
Golgi
tendon organs
the muscle tendons and transmit information about
tendon tension or rate of change of tension.
entirely or almost entirely for the purpose of intrinsic
amounts of information not only to the spinal cord but
also to the cerebellum and even to the cerebral cortex,
helping each of these portions of the nervous system
function to control muscle contraction.
Muscle Spindle
around 3 to 12 very small intrafusal muscle fibers that
are pointed at their ends and attached to the glycoca-
lyx of the surrounding large extrafusal skeletal muscle
Each intrafusal muscle fiber is a very small skeletal
its two ends—has few or no actin and myosin fila-
do contract are excited by small gamma motor nerve
fibers
neurons in the anterior horns of the spinal cord,
are also called
in contradistinc-
tion to the large alpha efferent fibers
nerve fibers) that innervate the extrafusal skeletal
readily see that the muscle spindle receptor can be
excited in two ways:
the primary ending and the
Primary Ending.
primary
ending or
and it transmits sensory signals to the spinal cord at a

stretched suddenly, excitation of the spindles causes
Whenever a muscle is
muscle stretch reflex.
The simplest manifestation of muscle spindle function
—that is, increased numbers of
firing. Thus, the spindles can send to the spinal cord
the muscle spindles increases the rate of firing,
emit sensory nerve impulses continuously. Stretching
degree of gamma nerve excitation, the muscle spindles
Normally, particularly when there is some
little influence on the dynamic response. Subsequent
chain fibers, enhances the static response while having
tion of the gamma-s fibers, which excite the nuclear
static response is hardly affected. Conversely, stimula-
spindle becomes tremendously enhanced, whereas the
nuclear bag fibers, the dynamic response of the muscle
intrafusal fibers. When the gamma-d fibers excite the
fibers, and the second excites mainly the nuclear chain
dynamic (gamma-d) and gamma-static (gamma-s).The
muscle spindle can be divided into two types: gamma-
The gamma motor nerves to the
Gamma Motor Nerves.
any change in length of the spindle receptor.
or negative, signals to the spinal cord to apprise it of
primary ending sends extremely strong, either positive
exactly opposite sensory signals occur. Thus, the
Conversely, when the spindle receptor shortens,
length stops increasing, this extra rate of impulse dis-
while the length is actually increasing.
large 17-micrometer sensory nerve fiber,
in a fraction of a second, the primary receptor trans-
only a fraction of a micrometer, if this increase occurs
rate of change
dynamic response,
especially powerfully. This excess stimulus of the
the spindle receptor increases suddenly, the primary
When the length of
ondary Ending) to Rate of Change of Receptor
of the spindle receptor,
mit these impulses for several minutes. This effect is
degree of stretching, and the endings continue to trans-
slowly,
When the receptor portion of the muscle
Endings to the Length of the Receptor—“Static”
tions are shown in Figure 54–3.
usually excited only by nuclear chain fibers. These rela-
the nuclear chain fibers. Conversely, the sec-
as shown by the bottom fiber in the figure. The primary
(three to nine), which are about half as large in diam-
nuclear chain fibers
top fiber in Figure 54–3, and (2)
central portion of the receptor area, as shown by the
nuclei are congregated in expanded “bags” in the
three in each spindle), in which several muscle fiber
intrafusal fibers: (1)
There are also two types of muscle spindle
that the type Ia fiber does, but often it spreads like
ending, as shown in Figures 54–2 and 54–3. This
nerve fiber in the entire body.
velocity of 70 to 120 m/sec, as rapidly as any type of
The Nervous System: C. Motor and Integrative Neurophysiology
676
Unit XI
Secondary Ending.
Usually one but sometimes two
smaller sensory nerve fibers—type II fibers with an
average diameter of 8 micrometers—innervate the
receptor region on one or both sides of the primary
sensory ending is called the secondary ending; some-
times it encircles the intrafusal fibers in the same way
branches on a bush.
Division of the Intrafusal Fibers into Nuclear Bag and Nuclear
Chain Fibers—Dynamic and Static Responses of the Muscle
Spindle.
nuclear bag muscle fibers (one to
eter and half as long as the nuclear bag fibers and have
nuclei aligned in a chain throughout the receptor area,
sensory nerve ending (the 17-micrometer sensory
fiber) is excited by both the nuclear bag intrafusal
fibers and
ondary ending (the 8-micrometer sensory fiber) is
Response of Both the Primary and the Secondary
Response.
spindle is stretched
the number of impulses
transmitted from both the primary and the secondary
endings increases almost directly in proportion to the
called the static response
meaning simply that both the primary and secondary
endings continue to transmit their signals for at least
several minutes if the muscle spindle itself remains
stretched.
Response of the Primary Ending (but Not the Sec-
Length—“Dynamic” Response.
ending (but not the secondary ending) is stimulated
primary ending is called the
which
means that the primary ending responds extremely
actively to a rapid
in spindle length.
Even when the length of a spindle receptor increases
mits tremendous numbers of excess impulses to the
but only
As soon as the
charge returns to the level of the much smaller static
response that is still present in the signal.
Control of Intensity of the Static and Dynamic Responses by the
first of these excites mainly the nuclear bag intrafusal
paragraphs illustrate that these two types of muscle
spindle responses are important in different types of
muscle control.
Continuous Discharge of the Muscle Spindles Under Normal
Conditions.
whereas shortening the spindle decreases the rate of
either positive signals
impulses to indicate stretch of a muscle—or negative
signals—below-normal numbers of impulses to indi-
cate that the muscle is unstretched.
Muscle Stretch Reflex
is the
Group II fiber
(secondary
efferent)
Nuclear bag fiber
(intrafusal muscle)
Nuclear chain fiber
(intrafusal muscle)
Plate
ending
Dynamic
g
fiber
(efferent)
Static
g
fiber
(efferent)
Trail ending
Group Ia fiber
(primary afferent)
control of movement. Physiol Rev 54:225, 1974.)
chain muscle spindle fibers. (Modified from Stein RB: Peripheral
Details of nerve connections from the nuclear bag and nuclear
Figure 54–3
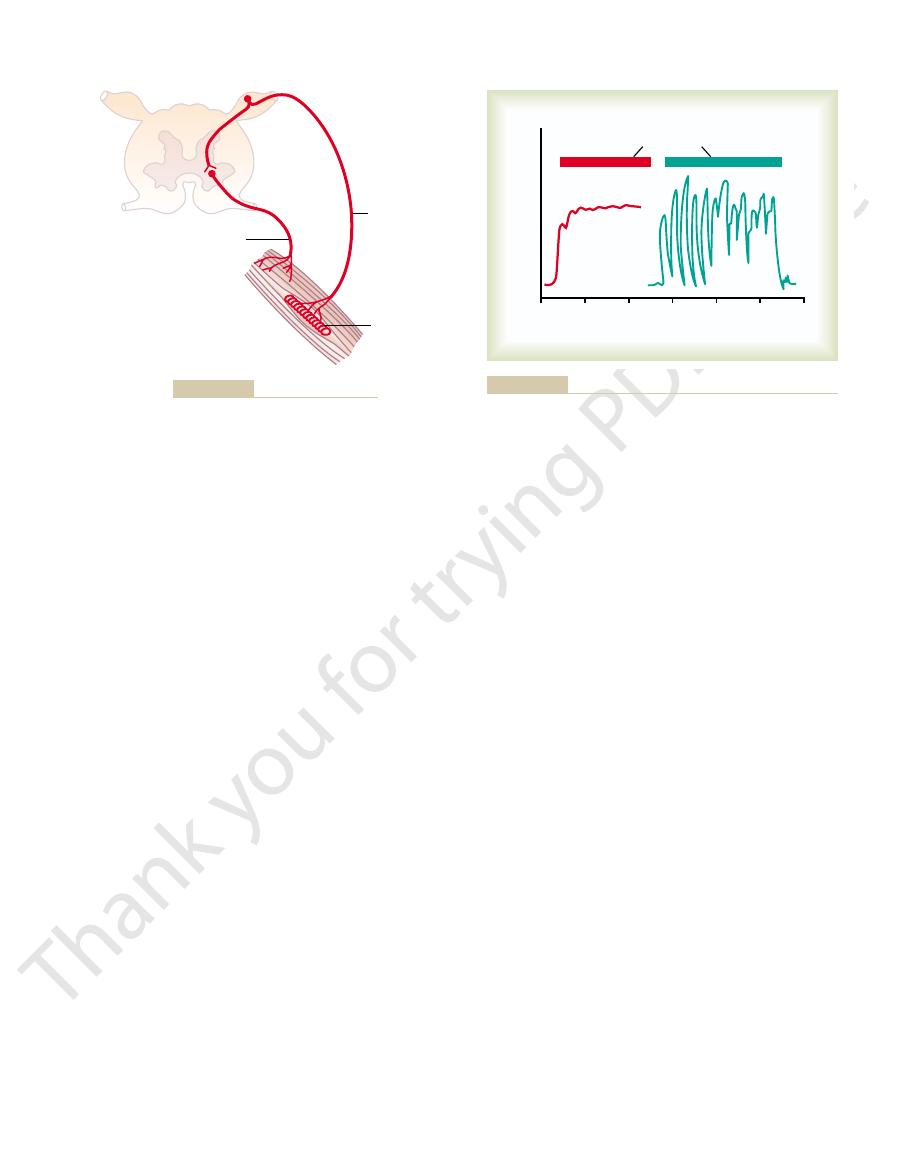
selves be jerky. This effect can also be called a
smooth muscle contractions, even though the primary
cally demonstrates the damping mechanism’s ability to
unsmooth muscle contraction. Thus, curve A graphi-
nerves had been sectioned 3 months earlier. Note the
signals per second. Curve B illustrates the same exper-
the muscle is excited at a slow frequency of only 8
is relatively smooth, even though the motor nerve to
the excited muscle is intact. Note that the contraction
Figure 54–5. In curve A, the muscle spindle reflex of
course of such a signal. This effect is demonstrated in
factorily, the muscle contraction is jerky during the
changing to another intensity level, and so forth. When
a few milliseconds, then decreasing in intensity, then
muscle in an unsmooth form, increasing in intensity for
, or smoothing, function,
movements. This is a
Stretch Reflexes
person’s nervous system specifically wills otherwise.
to remain reasonably constant, except when the
secondary endings. The importance of the static stretch
after. This reflex is elicited by the continuous static
stretch reflex
unstretched) to its new length, but then a weaker
The dynamic stretch reflex is over within a fraction
the reflex functions to oppose sudden changes in muscle
same muscle from which the signal originated. Thus,
to the spinal cord; this causes an instantaneous strong
stretched or unstretched, a strong signal is transmitted
or unstretch. That is, when a muscle is suddenly
endings of the muscle spindles, caused by rapid stretch
dynamic stretch reflex
The
stretch reflex can be divided into two components: the
The
serve other functions.
interneurons in the cord gray matter, and these trans-
muscle after excitation of the spindle. Most type II
monosynaptic pathway
which the muscle spindle fiber originated. Thus, this is
dorsal root of the spinal cord. A branch of this fiber
stretch reflex, showing a type Ia proprioceptor nerve
Figure 54–4
Neuronal Circuitry of the Stretch Reflex.
gistic muscles.
Motor Functions of the Spinal Cord; the Cord Reflexes
Chapter 54
677
reflex contraction of the large skeletal muscle fibers of
the stretched muscle and also of closely allied syner-
demonstrates the basic circuit of the muscle spindle
fiber originating in a muscle spindle and entering a
then goes directly to the anterior horn of the cord gray
matter and synapses with anterior motor neurons that
send motor nerve fibers back to the same muscle from
a
that allows a reflex signal to
return with the shortest possible time delay back to the
fibers from the muscle spindle terminate on multiple
mit delayed signals to the anterior motor neurons or
Dynamic Stretch Reflex and Static Stretch Reflexes.
dynamic stretch reflex and the static stretch reflex.
is elicited by the potent
dynamic signal transmitted from the primary sensory
reflex contraction (or decrease in contraction) of the
length.
of a second after the muscle has been stretched (or
static
continues for a prolonged period there-
receptor signals transmitted by both primary and
reflex is that it causes the degree of muscle contraction
“Damping” Function of the Dynamic and Static
An especially important function of the stretch reflex
is its ability to prevent oscillation or jerkiness of body
damping
as explained in the following paragraph.
Damping Mechanism in Smoothing Muscle Contraction.
Signals from the spinal cord are often transmitted to a
the muscle spindle apparatus is not functioning satis-
iment in an animal whose muscle spindle sensory
input signals to the muscle motor system may them-
signal
averaging function of the muscle spindle reflex.
Motor nerve
Stretch reflex
Muscle spindle
Sensory nerve
Neuronal circuit of the stretch reflex.
Figure 54–4
0
1
2
Stimulus
(8 per second)
Seconds
Force of contraction
B
A
3
sity Press, 1932.)
et al: Reflex Activity of the Spinal Cord. New York: Oxford Univer-
of the muscle spindle reflex in curve A. (Modified from Creed RS,
rior roots of the cord 82 days previously. Note the smoothing effect
whose muscle spindles were denervated by section of the poste-
ditions: curve A, in a normal muscle, and curve B, in a muscle
Muscle contraction caused by a spinal cord signal under two con-
Figure 54–5

the degree of facilitation of spinal cord centers. When
The muscle jerks are used by neurologists to assess
other words, sudden stretch of muscle spindles is all that
muscle or by striking the belly of the muscle itself. In
part of Figure 54–6 shows a myogram from the quadri-
that causes the lower leg to “jerk” forward. The upper
dynamic stretch reflex
with a reflex hammer; this instantaneously stretches the
elicit the knee jerk and other muscle jerks.The knee jerk
Clinically, a method used
spinal cord. This reflex is elicited as follows.
ground excitation, or “tone,” the brain is sending to the
reflexes. The purpose is to determine how much back-
ination on a patient, he or she elicits multiple stretch
Clinical Applications of the
procedures.
joints. This aids tremendously in performing the
tioning, excitation of the appropriate muscle spindles
opposing each other at the joint. The net effect is that
joint also increases, producing tight, tense muscles
sides of each joint are activated at the same time, reflex
their signal output. However, if the spindles on both
stretches the central receptor regions, thus increasing
spindles. This shortens the ends of the spindles and
tense motor action. To do this, the bulboreticular facil-
Position During Tense Action
during walking and running.
high density of muscle spindles, emphasis is given to
ticularly concerned with antigravity contractions, and
control of the gamma efferent system. However,
basal ganglia,
the brain stem and, secondarily, by impulses transmit-
The gamma efferent system is excited specifically by
Brain Areas for Control of the Gamma
be flail and sometimes be overstretched, in neither
tract and relax along with the large muscle fibers,
length. For instance, if the muscle spindle did not con-
the muscle spindle, regardless of any change in muscle
Second, it maintains the proper damping function of
contraction. Therefore, coactivation keeps the muscle
muscle fibers contract is twofold: First, it keeps the
The purpose of contracting the muscle spindle intra-
muscle fibers to contract at the same time.
motor neurons. This causes both the extrafusal skele-
gamma motor neurons are stimulated simultaneously,
brain to the alpha motor neurons, in most instances the
alpha motor fibers. Whenever signals are transmitted
A gamma efferent fibers rather than large type A
system, one needs to recognize that 31 per cent of all
To emphasize the importance of the gamma efferent
in Voluntary Motor Activity
Role of the Muscle Spindle
The Nervous System: C. Motor and Integrative Neurophysiology
678
Unit XI
the motor nerve fibers to the muscle are the small type
from the motor cortex or from any other area of the
an effect called coactivation of the alpha and gamma
tal muscle fibers and the muscle spindle intrafusal
fusal fibers at the same time that the large skeletal
length of the receptor portion of the muscle spindle
from changing during the course of the whole muscle
spindle reflex from opposing the muscle contraction.
the receptor portion of the spindle would sometimes
instance operating under optimal conditions for
spindle function.
Motor System
signals from the bulboreticular facilitatory region of
ted into the bulboreticular area from (1) the cerebel-
lum, (2) the
and (3) the cerebral cortex.
Little is known about the precise mechanisms of
because the bulboreticular facilitatory area is par-
because the antigravity muscles have an especially
the importance of the gamma efferent mechanism for
damping the movements of the different body parts
Muscle Spindle System Stabilizes Body
One of the most important functions of the muscle
spindle system is to stabilize body position during
itatory region and its allied areas of the brain stem
transmit excitatory signals through the gamma nerve
fibers to the intrafusal muscle fibers of the muscle
excitation of the skeletal muscles on both sides of the
the position of the joint becomes strongly stabilized,
and any force that tends to move the joint from its
current position is opposed by highly sensitized stretch
reflexes operating on both sides of the joint.
Any time a person must perform a muscle function
that requires a high degree of delicate and exact posi-
by signals from the bulboreticular facilitatory region
of the brain stem stabilizes the positions of the major
additional detailed voluntary movements (of fingers
or other body parts) required for intricate motor
Stretch Reflex
Almost every time a clinician performs a physical exam-
Knee Jerk and Other Muscle Jerks.
to determine the sensitivity of the stretch reflexes is to
can be elicited by simply striking the patellar tendon
quadriceps muscle and excites a
ceps muscle recorded during a knee jerk.
Similar reflexes can be obtained from almost any
muscle of the body either by striking the tendon of the
is required to elicit a dynamic stretch reflex.
0
200
400
Milliseconds
Muscle length
Patellar tendon struck
Knee jerk
Ankle clonus
600
800
and from the gastrocnemius muscle
Myograms recorded from the quadriceps muscle during elicita-
Figure 54–6
tion of the knee jerk (above)
during ankle clonus (below).

such destructive effects.
opposed by this negative reflex, can occasionally cause
ulation of muscles in the laboratory, which cannot be
bone. We know, for instance, that direct electrical stim-
taneous relaxation of the entire muscle. This effect is
tendon becomes extreme, the inhibitory effect from
When tension on the muscle and, therefore, on the
ment of too much tension on the muscle.
tive feedback
Thus, this reflex provides a
inhibitory.
cause reflex effects in the respective muscle. This reflex
muscle, signals are transmitted to the spinal cord to
When the Golgi tendon organs of a muscle tendon are
Its Importance
Inhibitory Nature of the Tendon Reflex and
without affecting adjacent muscles. The relation
This local circuit directly inhibits the individual muscle
cortex. The local cord signal excites a single
in a dorsal horn of the cord, through long fiber path-
both into local areas of the cord and, after synapsing
from the primary spindle endings, transmit signals
endings of the muscle spindle. These fibers, like those
ter, only slightly smaller than those from the primary
Ib nerve fibers that average 16 micrometers in diame-
are transmitted through large, rapidly conducting type
Central Nervous System.
Transmission of Impulses from the Tendon Organ into the
muscle.
response). Thus, Golgi tendon organs provide the
static response,
muscle spindle, has both a
The tendon organ, like the primary receptor of the
reflected by the tension in itself.
detects muscle length and changes in muscle length,
“tensed” by contracting or stretching the muscle. Thus,
nected to each Golgi tendon organ, and the organ is
pass. About 10 to 15 muscle fibers are usually con-
tendon organ, shown in Figure 54–7, is an encapsulated
The Golgi
Golgi Tendon Organ Helps Control Muscle Tension.
Golgi Tendon Re
ing force to it. If clonus occurs, the degree of facilitation
spinal cord, neurologists test patients for clonus by sud-
readily. To determine the degree of facilitation of the
stretch reflexes are highly facilitated, clonus develops
brain. For instance, in a decerebrate animal, in which the
clonus.
continues to oscillate, often for long periods; this is
this way, the stretch reflex of the gastrocnemius muscle
and the body falls once more to begin a new cycle. In
body, but this too dies out after a fraction of a second,
second time. Again, a dynamic stretch reflex lifts the
and the body falls again, thus stretching the spindles a
of a second, the reflex contraction of the muscle dies out
muscle, which lifts the body up again. After a fraction
cord. These impulses reflexively excite the stretched
the gastrocnemius muscles, stretch reflex impulses are
ankle clonus, as follows.
(see lower myogram, Figure 54–6). Oscil-
tions, the muscle jerks can oscillate, a phenomenon
of the body.
facilitatory area of the brain stem. Ordinarily, large
ably weakened or absent. These reflexes are used most
depressed or abrogated, the muscle jerks are consider-
gerated. Conversely, if the facilitatory impulses are
system into the cord, the muscle jerks are greatly exag-
Motor Functions of the Spinal Cord; the Cord Reflexes
Chapter 54
679
large numbers of facilitatory impulses are being trans-
mitted from the upper regions of the central nervous
frequently in determining the presence or absence of
muscle spasticity caused by lesions in the motor areas
of the brain or diseases that excite the bulboreticular
lesions in the motor areas of the cerebral cortex but not
in the lower motor control areas (especially lesions
caused by strokes or brain tumors) cause greatly exag-
gerated muscle jerks in the muscles on the opposite side
Clonus—Oscillation of Muscle Jerks.
Under some condi-
called clonus
lation can be explained particularly well in relation to
If a person standing on the tip ends of the feet sud-
denly drops his or her body downward and stretches
transmitted from the muscle spindles into the spinal
Clonus ordinarily occurs only when the stretch reflex
is highly sensitized by facilitatory impulses from the
denly stretching a muscle and applying a steady stretch-
is certain to be high.
flex
sensory receptor through which muscle tendon fibers
stimulated when this small bundle of muscle fibers is
the major difference in excitation of the Golgi tendon
organ versus the muscle spindle is that the spindle
whereas the tendon organ detects muscle tension as
dynamic response and a
responding intensely when the muscle
tension suddenly increases (the dynamic response) but
settling down within a fraction of a second to a lower
level of steady-state firing that is almost directly
proportional to the muscle tension (the static
nervous system with instantaneous information on the
degree of tension in each small segment of each
Signals from the tendon organ
ways such as the spinocerebellar tracts into the cere-
bellum and through still other tracts to the cerebral
inhibitory
interneuron that inhibits the anterior motor neuron.
between signals to the brain and function of the cere-
bellum and other parts of the brain for muscle control
is discussed in Chapter 56.
stimulated by increased tension in the connecting
is entirely
nega-
mechanism that prevents the develop-
the tendon organ can be so great that it leads to a
sudden reaction in the spinal cord that causes instan-
called the lengthening reaction; it is probably a pro-
tective mechanism to prevent tearing of the muscle or
avulsion of the tendon from its attachments to the
Tendon
Muscle
Nerve fiber (16
m
m)
Figure 54–7
Golgi tendon organ.

discharge may last for a second or more.
discharge, but after a strong pain stimulus, the after-
reflex; a weak tactile stimulus causes almost no after-
to occur. The duration of afterdischarge depends on
of afterdischarge, it takes many milliseconds for this
of the muscle returns toward the baseline, but because
cord. Finally, after the stimulus is over, the contraction
fatigue,
response appears. Then, in the next few seconds, the
after a pain nerve begins to be stimulated, the flexor
muscle during a flexor reflex.Within a few milliseconds
Figure 54–9 shows a typical myogram from a flexor
stimulus is over.
discharge
circuits to inhibit the antagonist muscles, called
the reflex to the necessary muscles for withdrawal; (2)
basic types of circuits: (1) diverging circuits to spread
pathway; however, most of the signals of the reflex tra-
The shortest possible circuit is a three- or four-neuron
neurons and only secondarily to the motor neurons.
The pathways for eliciting the flexor reflex do not
the hand from the painful stimulus.
of the upper arm become excited, thus withdrawing
lus is applied to the hand; as a result, the flexor muscles
for the flexor reflex. In this instance, a painful stimu-
portion of Figure 54–8 shows the neuronal pathways
The left-hand
withdrawal reflexes.
same type of reflex. Therefore, the many patterns of
fined to flexor muscles, even though it is basically the
drawn from the stimulus,
is painfully stimulated, that part will similarly be
a pinprick, heat, or a wound, for which reason it is also
powerfully by stimulation of pain endings, such as by
In its classic form, the flexor reflex is elicited most
object. This is called the
In the spinal or decerebrate animal, almost any type
that originate in all these areas.
Chapters 55 and 56, the information from these recep-
motor areas of the cerebral cortex. As discussed in
brain stem and, to a lesser extent, all the way to the
brain or spinal cord. Additional pathways transmit
120 m/sec, the most rapid conduction anywhere in the
For instance, the dorsal spinocerebellar tracts carry
of instantaneous changes taking place in the muscles.
cord control of motor function, these two sensory
with Motor Control from Higher Levels
Golgi Tendon Organs in Conjunction
Function of the Muscle Spindles and
inhibition. This spreads the muscle load over all the
reflex, whereas those that exert too little tension
forces of the separate muscle fibers.That is, those fibers
Possible Role of the Tendon Reflex to Equalize Contractile
The Nervous System: C. Motor and Integrative Neurophysiology
680
Unit XI
Force Among the Muscle Fibers.
Another likely function
of the Golgi tendon reflex is to equalize contractile
that exert excess tension become inhibited by the
become more excited because of absence of reflex
fibers and prevents damage in isolated areas of
a muscle where small numbers of fibers might be
overloaded.
of the Brain
Although we have emphasized the function of the
muscle spindles and Golgi tendon organs in spinal
organs also apprise the higher motor control centers
instantaneous information from both the muscle
spindles and the Golgi tendon organs directly to the
cerebellum at conduction velocities approaching
similar information into the reticular regions of the
tors is crucial for feedback control of motor signals
Flexor Reflex and the
Withdrawal Reflexes
of cutaneous sensory stimulus from a limb is likely to
cause the flexor muscles of the limb to contract,
thereby withdrawing the limb from the stimulating
flexor reflex.
called a nociceptive reflex, or simply a pain reflex. Stim-
ulation of touch receptors can also elicit a weaker and
less prolonged flexor reflex.
If some part of the body other than one of the limbs
with-
but the reflex may not be con-
these reflexes in the different areas of the body are
called
Neuronal Mechanism of the Flexor Reflex.
pass directly to the anterior motor neurons but instead
pass first into the spinal cord interneuron pool of
verse many more neurons and involve the following
recip-
rocal inhibition circuits; and (3) circuits to cause after-
lasting many fractions of a second after the
reflex begins to
which is characteristic of essen-
tially all complex integrative reflexes of the spinal
the intensity of the sensory stimulus that elicited the
FLEXOR
REFLEX
CROSSED EXTENSOR
REFLEX
Painful
stimulus
from hand
Polysynaptic
circuit
Excited
RECIPROCAL INHIBITION
Inhibited
Excited
Inhibited
Flexor reflex, crossed extensor reflex, and reciprocal inhibition.
Figure 54–8

This is the phenomenon of
often simultaneously inhibits the antagonist muscles.
instance, when a stretch reflex excites one muscle, it
associated with inhibition of another group. For
In the previous paragraphs, we pointed out several
Reciprocal Innervation
Reciprocal Inhibition and
cause the entire body to move away.
of the stimulus. The prolonged afterdischarge is of
This demonstrates the relatively long latency before
Figure 54–10 shows a typical myogram recorded
the interneuronal cells.
reflex. Again, it is presumed that this prolonged after-
removed, the crossed extensor reflex has an even
crossed extension. After the painful stimulus is
of the initial pain stimulus, it is certain that many
muscles. Because the crossed extensor reflex usually
right-hand portion of Figure 54–8 shows the neuronal
The
drawn limb.
This is called the
reflex in one limb, the opposite limb begins to extend.
Crossed Extensor Reflex
limbs because of their highly developed flexor reflexes.
any part of the body, it is especially applicable to the
principle, called the principle of “local sign,” applies to
away from the object causing the pain. Although this
outward. In other words, the integrative centers of
which sensory nerve is stimulated. Thus, a pain stimu-
The pattern of withdrawal that
from the painful stimulus.
over. During this time, other reflexes and actions of the
charge, the reflex can hold the irritated part away from
body from a stimulus. Further, because of afterdis-
Thus, the flexor reflex is appropriately organized to
sensory signal is over.
transmit impulses to the anterior motor neurons,
reverberating interneuron circuits. These, in turn,
after strong pain stimuli, almost certainly resulting
themselves. Also, prolonged afterdischarge occurs
charge, lasting for about 6 to 8 milliseconds, results
discharge circuits discussed in Chapter 46. Electro-
The afterdischarge that occurs in the flexor reflex
Motor Functions of the Spinal Cord; the Cord Reflexes
Chapter 54
681
almost certainly results from both types of repetitive
physiologic studies indicate that immediate afterdis-
from repetitive firing of the excited interneurons
from recurrent pathways that initiate oscillation in
sometimes for several seconds after the incoming
withdraw a pained or otherwise irritated part of the
the stimulus for 0.1 to 3 seconds after the irritation is
central nervous system can move the entire body away
Pattern of Withdrawal.
results when the flexor reflex is elicited depends on
lus on the inward side of the arm elicits not only
contraction of the flexor muscles of the arm but also
contraction of abductor muscles to pull the arm
the cord cause those muscles to contract that can
most effectively remove the pained part of the body
About 0.2 to 0.5 second after a stimulus elicits a flexor
crossed extensor reflex. Extension of
the opposite limb can push the entire body away from
the object causing the painful stimulus in the with-
Neuronal Mechanism of the Crossed Extensor Reflex.
circuit responsible for the crossed extensor reflex,
demonstrating that signals from sensory nerves cross
to the opposite side of the cord to excite extensor
does not begin until 200 to 500 milliseconds after onset
interneurons are involved in the circuit between the
incoming sensory neuron and the motor neurons of
the opposite side of the cord responsible for the
longer period of afterdischarge than does the flexor
discharge results from reverberating circuits among
from a muscle involved in a crossed extensor reflex.
the reflex begins and the long afterdischarge at the end
benefit in holding the pained area of the body away
from the painful object until other nervous reactions
times that excitation of one group of muscles is often
reciprocal inhibition, and
the neuronal circuit that causes this reciprocal relation
Seconds
Flexor contraction
0
1
Duration of stimulus
Fatigue
Afterdischarge
2
3
stimulus is over.
an interval of fatigue, and, finally, afterdischarge after the input
Myogram of the flexor reflex showing rapid onset of the reflex,
Figure 54–9
1
2
3
4
Extensor contraction
Seconds
0
Duration of
stimulus
Afterdischarge
longed afterdischarge.
Myogram of a crossed extensor reflex showing slow onset but pro-
Figure 54–10

another manifestation of reciprocal innervation, this
the forelimbs and hindlimbs. This diagonal response is
limbs. In general, stepping occurs diagonally between
shown in Figure 54–12, the stretch on the limbs occa-
up from the floor and its legs are allowed to dangle, as
Diagonal Stepping of All Four Limbs—“Mark Time” Reflex.
from reciprocal innervation between the two limbs.
site limb ordinarily moves backward.This effect results
occurs in the forward direction in one limb, the oppo-
cord is not split down its center, every time stepping
controller.
Thus, the cord is an intelligent walking
forward to be placed over the obstruction. This is the
sequence, the foot will be lifted higher and proceed
the forward thrust will stop temporarily; then, in rapid
be even more complex. For instance, if the top of the
fact, the cord mechanism for control of stepping can
when the foot is allowed to walk along a surface. In
controlling foot pressure and frequency of stepping
The sensory signals from the footpads and from the
neurons controlling agonist and antagonist muscles.
matrix of the cord itself, oscillating between the
nerves have been cut, and it seems to result mainly
This oscillation back and forth between flexor and
repeated over and over.
extension. Then flexion occurs again, and the cycle is
individual stepping functions. Forward flexion of the
between the two limbs, each hindlimb can still perform
limbs of spinal animals. Indeed, even when the lumbar
Stepping and Walking Movements
forelimbs.
of the thoracic cord, the walking movements of
walk using its hindlimbs in addition to its forelimbs.
posture are integrated in the spinal cord. Indeed, an
cord righting reflex.
raise itself to the standing position. This is called
its side, it will make incoordinate movements trying to
When a spinal animal is laid on
animal from falling to that side.
This helps keep an
sure on one side causes extension in that direction, an
mines the direction in which the limb will extend; pres-
The locus of the pressure on the pad of the foot deter-
responsible for the flexor and cross extensor reflexes.
The positive supportive reaction involves a complex
support the weight of the body. This reflex is called the
feet, the reflex often stiffens the limbs sufficiently to
been transected for several months—that is, after the
the pressure applied to the foot. Indeed, this reflex
Reflexes of Posture and
the original reflex to reassume its previous intensity.
flexion. Finally, removal of the stronger reflex allows
body. This stronger reflex sends reciprocal inhibitory
while this reflex is still being elicited, a stronger flexor
inhibition. In this instance, a moderate but prolonged
Figure 54–11 shows a typical example of reciprocal
extensor muscle reflexes described earlier.
sides of the body, as exemplified by the flexor and
Likewise, reciprocal
The Nervous System: C. Motor and Integrative Neurophysiology
682
Unit XI
is called reciprocal innervation.
relations often exist between the muscles on the two
flexor reflex is elicited from one limb of the body;
reflex is elicited in the limb on the opposite side of the
signals to the first limb and depresses its degree of
Locomotion
Postural and Locomotive Reflexes
of the Cord
Positive Supportive Reaction.
Pressure on the footpad of
a decerebrate animal causes the limb to extend against
is so strong that if an animal whose spinal cord has
reflexes have become exaggerated—is placed on its
positive supportive reaction.
circuit in the interneurons similar to the circuits
effect called the magnet reaction.
Cord “Righting” Reflexes.
the
Such a reflex demonstrates
that some relatively complex reflexes associated with
animal with a well-healed transected thoracic cord
between the levels for forelimb and hindlimb innerva-
tion can right itself from the lying position and even
In the case of an opossum with a similar transection
the hindlimbs are hardly different from those in a
normal opossum—except that the hindlimb walking
movements are not synchronized with those of the
Rhythmical Stepping Movements of a Single Limb.
Rhythmi-
cal stepping movements are frequently observed in the
portion of the spinal cord is separated from the
remainder of the cord and a longitudinal section is
made down the center of the cord to block neuronal
connections between the two sides of the cord and
limb is followed a second or so later by backward
extensor muscles can occur even after the sensory
from mutually reciprocal inhibition circuits within the
position sensors around the joints play a strong role in
foot encounters an obstruction during forward thrust,
stumble reflex.
Reciprocal Stepping of Opposite Limbs.
If the lumbar spinal
If
a well-healed spinal animal (with spinal transection in
the neck above the forelimb area of the cord) is held
sionally elicits stepping reflexes that involve all four
Flexor contraction
0
1
Duration of inhibitory stimulus
Duration of flexor reflex stimulus
2
3
Seconds
site side of the body.
by an inhibitory stimulus from a stronger flexor reflex on the oppo-
Myogram of a flexor reflex showing reciprocal inhibition caused
Figure 54–11

tion, all the segmental reflexes can at times be elicited
(see Chapter 31) or the colon (see Chapter 63). In addi-
response to peritoneal irritation (see Chapter 66); and
functions of the gut (see Chapter 62); (4) peritoneoin-
heat on the surface of the body (see Chapter 73); (3)
Chapter 73); (2) sweating, which results from localized
other chapters. Briefly, these include (1) changes in vas-
grated in the spinal cord, most of which are discussed in
Many types of segmental autonomic reflexes are inte-
Spinal Cord
blown muscle cramp ensues.
itive feedback develops, so that a small amount of initial
cord to increase the intensity of contraction. Thus, pos-
sensory receptors even more, which causes the spinal
The contraction is believed to stimulate the same
flow, or overexercise, can elicit pain or other sensory
mality of a muscle, such as severe cold, lack of blood
follows: Any local irritating factor or metabolic abnor-
typical muscle cramp. Electromyographic studies indi-
abdominal operations.
this reason, deep anesthesia is usually required for intra-
extruding the intestines through the surgical wound. For
abdominal muscles to contract extensively, sometimes
for instance, during abdominal operations, pain im-
peritonitis allows the spastic muscle to relax. The same
peritonitis. Here again, relief of the pain caused by the
priate positions.
as ether anesthesia, also relieves the spasm. One of
spasm; a deep general anesthetic of the entire body, such
tonically. Pain relief obtained by injecting a local anes-
initiated from the broken edges of the bone, which
round a broken bone. This results from pain impulses
In many, if not most, instances, localized pain is the
In human beings, local muscle spasm is often observed.
Spinal Cord Reflexes That
ments of locomotion, involves reciprocal innervation
to-and-fro movement,
The
crosses the midline, the first paw stops scratching and
make the reflex even more complicated, when the flea
to bring the paw to the position of the crawling flea. To
find its position, even though 19 muscles in the limb
as the shoulder of a spinal animal, the hind paw can still
developed function. If a flea is crawling as far forward
The
movement.
to-and-fro scratching
the surface of the body, and (2) a
It involves two functions: (1) a
tickle sen-
itch
the scratch reflex, which is initiated by
poses the animal to keep galloping and, therefore, con-
hindlimbs are stimulated about equally; this predis-
ground during galloping, both forelimbs and both
walking. Conversely, when the animal strikes the
ulated, which would predispose the animal to continue
terns of walking and galloping, because in walking,
walking reflex. This is in keeping with the normal pat-
same time; unequal stimulation elicits the diagonal
while both hindlimbs move forward. This often occurs
cord between the forelimbs and hindlimbs. Such a
Motor Functions of the Spinal Cord; the Cord Reflexes
Chapter 54
683
time occurring the entire distance up and down the
walking pattern is called a mark time reflex.
Galloping Reflex.
Another type of reflex that occasion-
ally develops in a spinal animal is the galloping reflex,
in which both forelimbs move backward in unison
when almost equal stretch or pressure stimuli are
applied to the limbs on both sides of the body at the
only one forelimb and one hindlimb at a time are stim-
tinues this pattern of motion.
Scratch Reflex
An especially important cord reflex in some animals is
or
sation.
position sense that
allows the paw to find the exact point of irritation on
position sense of the scratch reflex is a highly
must be contracted simultaneously in a precise pattern
the opposite paw begins the to-and-fro motion and
eventually finds the flea.
like the stepping move-
circuits that cause oscillation.
Cause Muscle Spasm
cause of the local spasm.
Muscle Spasm Resulting from a Broken Bone.
One type of
clinically important spasm occurs in muscles that sur-
cause the muscles that surround the area to contract
thetic at the broken edges of the bone relieves the
these two anesthetic procedures is often necessary
before the spasm can be overcome sufficiently for the
two ends of the bone to be set back into their appro-
Abdominal Muscle Spasm in Peritonitis.
Another type of
local spasm caused by cord reflexes is abdominal spasm
resulting from irritation of the parietal peritoneum by
type of spasm often occurs during surgical operations;
pulses from the parietal peritoneum often cause the
Muscle Cramps.
Still another type of local spasm is the
cate that the cause of at least some muscle cramps is as
signals transmitted from the muscle to the spinal cord,
which in turn cause reflex feedback muscle contraction.
irritation causes more and more contraction until a full-
Autonomic Reflexes in the
cular tone resulting from changes in local skin heat (see
intestinointestinal reflexes that control some motor
testinal reflexes that inhibit gastrointestinal motility in
(5) evacuation reflexes for emptying the full bladder
simultaneously in the form of the so-called mass reflex,
described next.
Figure 54–12
Diagonal stepping movements exhibited by a spinal animal.

Res Brain Res Rev 40:257, 2002.
locomotion in the cat following spinal cord lesions. Brain
Rossignol S, Bouyer L, Barthelemy D, et al: Recovery of
motoneuronal excitability. Physiol Rev 80:767, 2000.
Rekling JC, Funk GD, Bayliss DA, et al: Synaptic control of
feedback. Prog Brain Res 143:123, 2004.
Pearson KG: Generating the walking gait: role of sensory
homeostasis. Neuron 37:2, 2003.
Marder E, Prinz AA: Current compensation in neuronal
control of rhythmic movements. Curr Biol 11:R986, 2001.
Marder E, Bucher D: Central pattern generators and the
Science, 4th ed. New York: McGraw-Hill, 2000.
Kandel ER, Schwartz JH, Jessell TM: Principles of Neural
Rev 40:19, 2002.
Jankowska E, Hammar I: Spinal interneurons: how can
mammals. J Physiol 533:31, 2001.
Jankowska E: Spinal interneuronal systems: identification,
interneurons. Curr Opin Neurobiol 13:42, 2003.
Helms AW, Johnson JE: Specification of dorsal spinal cord
Churchill Livingstone, 2003.
Haines DE, Lancon JA: Review of Neuroscience. New York:
Churchill Livingstone, 1997.
Haines DE: Fundamental Neuroscience. New York:
their place. Curr Biol 14:R166, 2004.
Guthrie S: Neuronal development: putting motor neurons in
muscles of the withdrawal reflex. Trends Neurosci 27:169,
Grillner S: Muscle twitches during sleep shape the precise
neuronal networks. Nat Rev Neurosci 4:573, 2003.
Grillner S: The motor infrastructure: from ion channels to
spinal cord. Physiol Rev 80:615, 2000.
Glover JC: Development of specific connectivity between
Brain Res Brain Res Rev 40:152, 2002.
Garwicz M: Spinal reflexes provide motor error signals to
80:83, 2000.
in gait and posture: comparative aspects. Physiol Rev
Duysens J, Clarac F, Cruse H: Load-regulating mechanisms
in therapy of spinal cord injury. Annu Rev Med 55:255,
Dobkin BH, Havton LA: Basic advances and new avenues
Neurophysiol 114:1379, 2003.
Dietz V: Spinal cord pattern generators for locomotion. Clin
Neurosci 3:781, 2002.
Dietz V: Proprioception and locomotor disorders. Nat Rev
Neurobiol 13:96, 2003.
of the monosynaptic stretch reflex circuit. Curr Opin
Chen HH, Hippenmeyer S, Arber S, Frank E: Development
tions. News Physiol Sci 19:85, 2004.
Buffelli M, Busetto G, Bidoia C, et al: Activity-dependent
cases they eventually return. These effects are
first few weeks after cord transection, but in most
3. The sacral reflexes for control of bladder and colon
stepping reflexes.
postural antigravity reflexes, and remnants of
progressively more complex reflexes: flexor reflexes,
stretch reflexes, followed in order by the
transected. The first reflexes to return are the
hyperexcitable, particularly if a few facilitatory
humans, some reflexes may eventually become
sometimes required. In both animals and
in human beings, 2 weeks to several months are
In lower animals, a few hours to a few days are
2. All skeletal muscle reflexes integrated in the spinal
in human beings.
ordinarily returns to normal within a few days, even
blocked almost to extinction. The pressure
1. At onset of spinal shock, the arterial blood
functions.
complete; conversely, sometimes recovery is excessive,
to a day or so, but in human beings, the return is often
the loss. In most nonprimates, excitability of the cord
facilitatory impulses, they increase their own natural
nervous system—that is, after they lose their source of
gradually regain their excitability. This seems to be a
After a few hours to a few weeks, the spinal neurons
tracts, and corticospinal tracts.
mitted through the reticulospinal tracts, vestibulospinal
cord from higher centers, particularly discharge trans-
The reason for this is that normal activity of the cord
spinal shock.
the point of total silence, a reaction called
ing the cord reflexes, immediately become depressed to
upper neck, at first, essentially all cord functions, includ-
When the spinal cord is suddenly transected in the
Spinal Cord Transection and
seizures, which involve reverberating circuits that occur
at once. This is similar to the mechanism of epileptic
Because the mass reflex can last for minutes, it pre-
areas of the body break out into profuse sweating.
to a systolic pressure well over 200 mm Hg; and (4) large
rial pressure often rises to maximal values, sometimes
colon and bladder are likely to evacuate; (3) the arte-
skeletal muscles goes into strong flexor spasm; (2) the
cord. The effects are (1) a major portion of the body’s
of the type of stimulus, the resulting reflex, called the
as overdistention of the bladder or the gut. Regardless
stimulus to the skin or excessive filling of a viscus, such
cord. The usual stimulus that causes this is a strong pain
active, causing massive discharge in large portions of the
In a spinal animal or human being, some-
The Nervous System: C. Motor and Integrative Neurophysiology
684
Unit XI
Mass Reflex.
times the spinal cord suddenly becomes excessively
mass reflex, involves large portions or even all of the
sumably results from activation of great numbers of
reverberating circuits that excite large areas of the cord
in the brain instead of in the cord.
Spinal Shock
neurons depends to a great extent on continual tonic
excitation by the discharge of nerve fibers entering the
natural characteristic of neurons everywhere in the
degree of excitability to make up at least partially for
centers returns essentially to normal within a few hours
delayed for several weeks and occasionally is never
with resultant hyperexcitability of some or all cord
Some of the spinal functions specifically affected
during or after spinal shock are the following:
pressure falls instantly and drastically—sometimes
to as low as 40 mm Hg—thus demonstrating that
sympathetic nervous system activity becomes
cord are blocked during the initial stages of shock.
required for these reflexes to return to normal;
pathways remain intact between the brain and the
cord while the remainder of the spinal cord is
evacuation are suppressed in human beings for the
discussed in Chapters 31 and 66.
References
synaptic competition at mammalian neuromuscular junc-
2004.
cerebellar modules—relevance for motor coordination.
premotor neurons and motoneurons in the brain stem and
2004.
multifunctional character and reconfigurations in
studies in animals contribute to the understanding of
spinal interneuronal systems in man? Brain Res Brain Res