
allows utilization of most of the energy in the incoming sound waves.
perfect for sound frequencies between 300 and 3000 cycles per second, which
of the cochlea. Indeed, the impedance matching is about 50 to 75 per cent of
matching
Therefore, the tympanic membrane and ossicular system provide
membrane. Because fluid has far greater inertia than air does, it is easily under-
averages 3.2 square millimeters. This 17-fold difference times the 1.3-fold ratio
brane is about 55 square millimeters, whereas the surface area of the stapes
movement about 1.3 times. In addition, the surface area of the tympanic mem-
Instead, the system actually reduces the distance but increases the
does not increase the movement distance of the stapes, as is commonly believed.
amplitude of the handle of the malleus. Therefore, the ossicular lever system
The amplitude of movement of the
time the tympanic membrane moves inward, and to pull backward on the fluid
The articulation of the incus with the stapes causes the stapes to push forward
imately at the border of the tympanic membrane.
the combined malleus and incus act as a single lever, having its fulcrum approx-
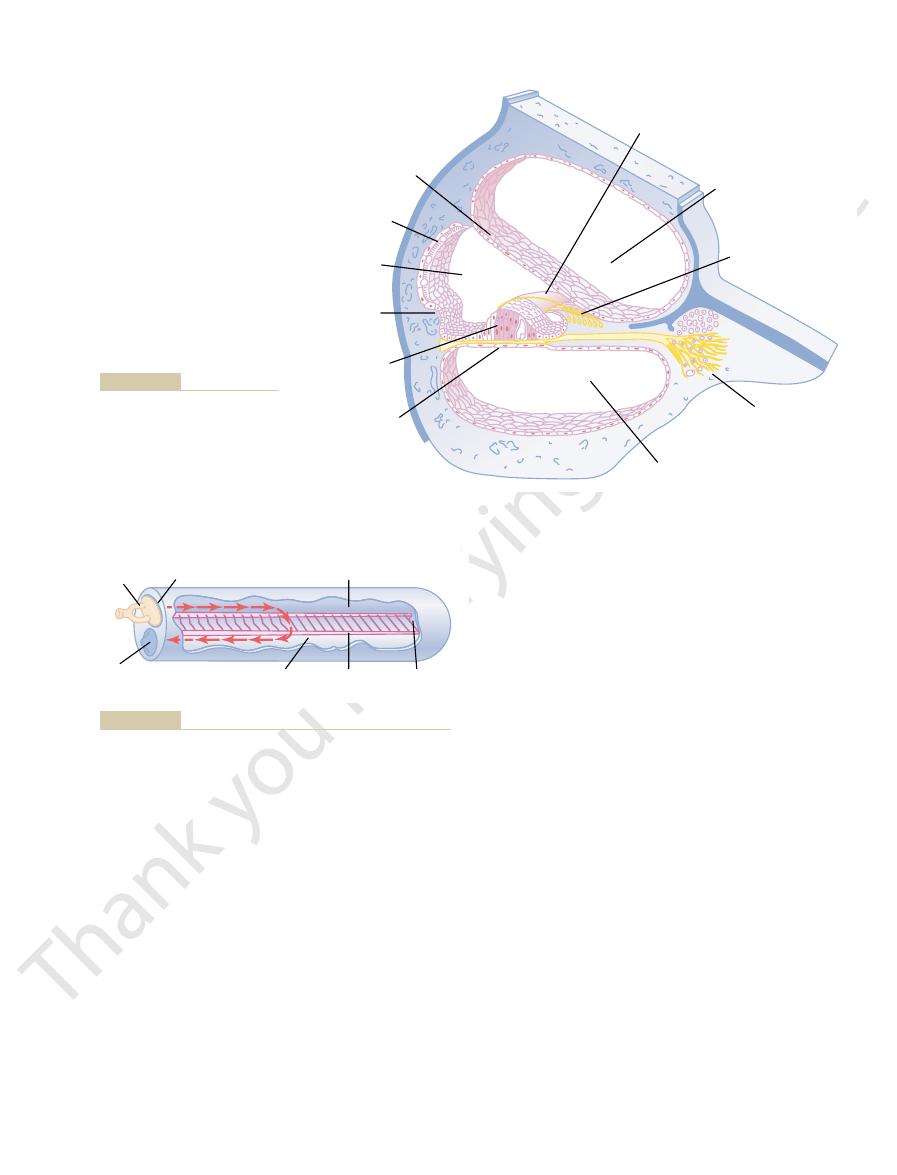
The ossicles of the middle ear are suspended by ligaments in such a way that
ossicles, which would not be true if the membrane were lax.
which keeps the tympanic membrane tensed. This allows sound
tympani muscle,
panic membrane, and this point of attachment is constantly pulled by the
The tip end of the handle of the malleus is attached to the center of the tym-
oval window
membranous labyrinth
stapes,
ments, so that whenever the malleus moves, the incus moves with it. The oppo-
The malleus is bound to the
malleus.
(the inner ear). Attached to the tympanic membrane
cochlea
ossicles,
eardrum
Figure 52–1 shows the
to the Cochlea
Conduction of Sound from the Tympanic Membrane
Tympanic Membrane and the Ossicular System
the central nervous system, where its meaning is
quencies, and transmits auditory information into
ear receives sound waves, discriminates their fre-
This chapter describes the mechanisms by which the
The Sense of Hearing
C
H
A
P
T
E
R
5
2
651
deciphered.
tympanic membrane (commonly called the
) and
the
which conduct sound from the tympanic membrane through the
middle ear to the
is the handle of the
incus by minute liga-
site end of the incus articulates with the stem of the
and the faceplate of
the stapes lies against the
of the cochlea in the opening
of the
.
tensor
vibrations on any portion of the tympanic membrane to be transmitted to the
on the oval window and on the cochlear fluid on the other side of window every
every time the malleus moves outward.
“Impedance Matching” by the Ossicular System.
stapes faceplate with each sound vibration is only three fourths as much as the
force of
of the lever system causes about 22 times as much total force to be exerted on
the fluid of the cochlea as is exerted by the sound waves against the tympanic
stood that increased amounts of force are needed to cause vibration in the fluid.
impedance
between the sound waves in air and the sound vibrations in the fluid

First, note that Reissner’s membrane is missing from
uncoiled cochlea for conduction of sound vibrations.
Figure 52–4 diagrams the functional parts of the
vibrations.
They are the receptive end organs
hair cells.
cells, the
basilar membrane.
), shown in Figure 52–3;
Reissner’s membrane
The scala vestibuli and scala media are sepa-
52–3. It consists of three tubes coiled side by side: (1)
Figure 52–1 and in cross section in Figures 52–2 and
The cochlea is a system of coiled tubes, shown in
Functional Anatomy of the Cochlea
to the bone.
the person to hear the sound. However, the energy
especially on the mastoid process near the ear, causes
tor placed on any bony protuberance of the skull, but
priate conditions, a tuning fork or an electronic vibra-
vibrations in the cochlea itself.Therefore, under appro-
labyrinth,
bony cavity in the temporal bone, called the
cochlea,
Because the inner ear, the
Transmission of Sound Through Bone
sensitivity to his or her own speech. This effect is acti-
stapedius muscles is to decrease a person’s hearing
second, where most of the pertinent information
concentrate on sounds above 1000 cycles per
environments. This usually removes a major share
low-frequency sounds in loud
2. To
1. To
between a loud voice and a whisper. The function of
bels, which is about the same difference as that
lower-frequency sound transmission by 30 to 40 deci-
This
cycles per second.
frequency sound, mainly frequencies below 1000
ossicular system to develop increased rigidity, thus
stapedius muscle pulls the stapes outward. These two
The tensor tympani muscle
tensor tympani muscle.
and, to a lesser extent, the
central nervous system, a reflex occurs after a latent
When loud sounds are transmitted
Attenuation of Sound by Contraction of the Tensor Tympani and
at the oval window. However, the sensitivity for
membrane, sound waves can still travel directly
The Nervous System: B. The Special Senses
652
Unit X
In the absence of the ossicular system and tympanic
through the air of the middle ear and enter the cochlea
hearing is then 15 to 20 decibels less than for ossicu-
lar transmission—equivalent to a decrease from a
medium to a barely perceptible voice level.
Stapedius Muscles.
through the ossicular system and from there into the
period of only 40 to 80 milliseconds to cause contrac-
tion of the stapedius muscle
pulls the handle of the malleus inward while the
forces oppose each other and thereby cause the entire
greatly reducing the ossicular conduction of low-
attenuation reflex can reduce the intensity of
this mechanism is believed to be twofold:
protect the cochlea from damaging vibrations
caused by excessively loud sound.
mask
of the background noise and allows a person to
in voice communication is transmitted.
Another function of the tensor tympani and
vated by collateral nerve signals transmitted to these
muscles at the same time that the brain activates the
voice mechanism.
is embedded in a
bony
vibrations of the entire skull can cause fluid
available even in loud sound in the air is not sufficient
to cause hearing via bone conduction unless a special
electromechanical sound-amplifying device is applied
Cochlea
the scala vestibuli, (2) the scala media, and (3) the scala
tympani.
rated from each other by
(also
called the vestibular membrane
the scala tympani and scala media are separated from
each other by the
On the surface of
the basilar membrane lies the organ of Corti, which
contains a series of electromechanically sensitive
that generate nerve impulses in response to sound
Cochlear
nerve
Spiral
ganglion
Cochlea
Oval window
Scala tympani
Stapes
Incus
Malleus
Scala vestibuli
Round
window
Auditory canal
Tympanic membrane
inner ear.
Tympanic membrane, ossicular system of the middle ear, and
Figure 52–1
Basilar
membrane
Spiral organ
of Corti
Vestibular membrane
Scala vestibuli
Stria
vascularis
Scala media
Scala tympani
Cochlear nerve
Spiral ganglion
Spiral
ligament
of the Human Body. Philadelphia: Lea & Febiger, 1948.)
Cochlea. (Redrawn from Gray H, Goss CM [eds]: Gray’s Anatomy
Figure 52–2
must vibrate along the cochlear tubules.
of increased “loading” with extra masses of fluid that
low-
enter the cochlea through the oval window. But
brane occurs near the base, where the sound waves
Thus,
best at a low frequency.
long, limber fibers near the tip of the cochlea vibrate
cochlea vibrate best at a very high frequency, while the
result, the stiff, short fibers near the oval window of the
overall stiffness decreases more than 100-fold. As a
the oval window to the helicotrema, so that their
of the fibers, however,
The
cochlea (the “helicotrema”), a 12-fold increase in
the base of the cochlea to the apex, increasing from a
The
one end, they can vibrate like the reeds of a
membrane. Because the fibers are stiff and free at
olus) but are not fixed at their distal ends, except
toward the outer wall. These fibers are stiff, elastic,
modiolus,
from the bony center of the cochlea, the
contains 20,000 to 30,000
rates the scala media from the scala tympani. It
The
Basilar Membrane and Resonance in the Cochlea.
forward in the scala vestibuli and scala media, and
tions. Inward movement causes the fluid to move
window’s edges by a loose annular ligament so that it
faceplate of the stapes at the oval window. The face-
later in the chapter.)
function of the sound-receptive hair cells, as discussed
Reissner’s membrane is to maintain a special kind of
sidered to be a single chamber. (The importance of
cerned, the scala vestibuli and scala media are con-
Therefore, as far as fluid conduction of sound is con-
this figure. This membrane is so thin and so easily
Chapter 52
The Sense of Hearing
653
moved that it does not obstruct the passage of sound
vibrations from the scala vestibuli into the scala media.
fluid in the scala media that is required for normal
Sound vibrations enter the scala vestibuli from the
plate covers this window and is connected with the
can move inward and outward with the sound vibra-
outward movement causes the fluid to move back-
ward.
basilar membrane is a fibrous membrane that sepa-
basilar fibers that project
reedlike structures that are fixed at their basal ends
in the central bony structure of the cochlea (the modi-
that the distal ends are embedded in the loose basilar
harmonica.
lengths of the basilar fibers increase progres-
sively beginning at the oval window and going from
length of about 0.04 millimeter near the oval and
round windows to 0.5 millimeter at the tip of the
length.
diameters
decrease from
high-frequency resonance of the basilar mem-
frequency resonance occurs near the helicotrema,
mainly because of the less stiff fibers but also because
Scala vestibuli
Tectorial membrane
Scala tympani
Basilar membrane
Organ of Corti
Spiral
prominence
Spiral limbus
Reissner's membrane
Spiral ganglion
Scala media
Stria vascularis
Histology, 11th ed. Philadelphia:
Bloom & Fawcett: A Textbook of
Colard Keene. From Fawcett DW:
Section through one of the turns
Figure 52–3
of the cochlea. (Drawn by Sylvia
WB Saunders, 1986.)
Scala vestibuli
and scala media
Oval
window
Basilar
membrane
Scala
tympani
Round
window
Stapes
Helicotrema
Movement of fluid in the cochlea after forward thrust of the stapes.
Figure 52–4
tion for different frequencies, demonstrating that the
Figure 52–6
frequency.
cycle. This is the
point but is moving inward. The shaded area around
outward, and (d) has moved back again to the neutral
moved back to the neutral point, (c) is all the way
when the stapes (a) is all the way inward, (b) has
The dashed curves of Figure 52–6
Amplitude Pattern of Vibration of the Basilar Mem-
be discriminated from one another.
the basilar membrane, and their frequencies could not
Without this, all the high-frequency waves would be
separate from one another on the basilar membrane.
mission of the wave allows the high-frequency sounds
farther along the membrane. This rapid initial trans-
farther into the cochlea. The cause of this is the high
travels the entire distance along the membrane.
and then dies, and a very low frequency sound wave
medium-frequency sound wave travels about halfway
before it reaches its resonant point and dies, a
membrane. Thus, a high-frequency sound wave travels
sipated. Consequently, the wave dies at this point and
this point, the basilar membrane can vibrate back and
quency equal to the respective sound frequency. At
frequencies. Each wave is relatively weak at the outset
Note in Figure 52–5 the different
rial walls, which is discussed in Chapter 15; it is also
a very low frequency wave. Movement
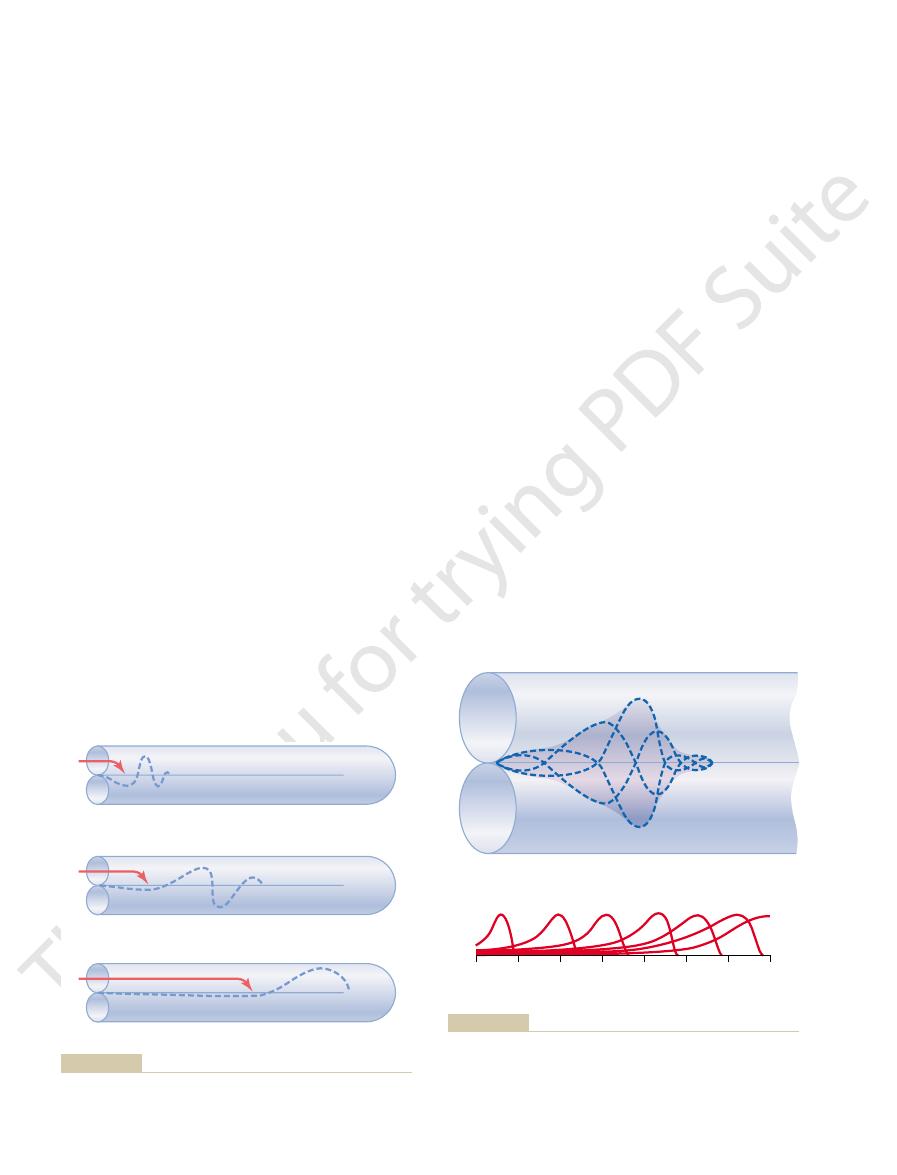
Figure 52–5
a medium-frequency wave; and
brane; Figure 52–5
ment of a high-frequency wave down the basilar mem-
as shown in Figure 52–5. Figure 52–5
round window initiates a fluid wave that “travels”
round window. However, the elastic tension that is
walls. The initial effect of a sound wave entering at the
window, the
oval
When the foot of the stapes moves inward against the
Cochlea—“Traveling Wave”
Transmission of Sound Waves in the
The Nervous System: B. The Special Senses
654
Unit X
round window must bulge outward
because the cochlea is bounded on all sides by bony
oval window is to cause the basilar membrane at
the base of the cochlea to bend in the direction of the
built up in the basilar fibers as they bend toward the
along the basilar membrane toward the helicotrema,
A shows move-
B,
C,
of the wave along the basilar membrane is compara-
ble to the movement of a pressure wave along the arte-
comparable to a wave that travels along the surface of
a pond.
Pattern of Vibration of the Basilar Membrane for Different
Sound Frequencies.
patterns of transmission for sound waves of different
but becomes strong when it reaches that portion of the
basilar membrane that has a natural resonant fre-
forth with such ease that the energy in the wave is dis-
fails to travel the remaining distance along the basilar
only a short distance along the basilar membrane
Another feature of the traveling wave is that it
travels fast along the initial portion of the basilar
membrane but becomes progressively slower as it goes
coefficient of elasticity of the basilar fibers near the
oval window and a progressively decreasing coefficient
to travel far enough into the cochlea to spread out and
bunched together within the first millimeter or so of
brane.
A show the
position of a sound wave on the basilar membrane
these different waves shows the extent of vibration of
the basilar membrane during a complete vibratory
amplitude pattern of vibration of
the basilar membrane for this particular sound
B shows the amplitude patterns of vibra-
High frequency
A
B
C
Medium frequency
Low frequency
medium-, and low-frequency sounds.
“Traveling waves” along the basilar membrane for high-,
Figure 52–5
15
25
10
20
30
4000
2000
1000
600
400
200
A
B
Distance from stapes (millimeters)
0
5
8000
a
b
d
c
Frequency
35
different frequencies.
frequencies between 200 and 8000 cycles per second, showing
Amplitude patterns for sounds of
a medium-frequency sound.
Amplitude pattern of vibration of the basilar membrane for
Figure 52–6
A,
B,
the points of maximum amplitude on the basilar membrane for the

of the inner hair cells at different sound pitches, a
loss occurs. Therefore, it has been proposed that the
remain fully functional, a large amount of hearing
cells rather than by the outer cells. Yet, despite this, if
outer hair cells as inner hair cells, about 90 per cent of
Auditory Signals Are Transmitted Mainly by the Inner Hair Cells.
whenever the basilar membrane vibrates.
the tectorial membrane. Thus, the hair cells are excited
The inward and outward motion causes the
outward.
ward, the reticular lamina rocks downward and
olus. Then, when the basilar membrane moves down-
inward
basilar fibers, the rods of Corti, and the reticular
which are attached tightly to the basilar fibers. The
rigid structure composed of a flat plate, called the
The outer ends of the hair cells are fixed tightly in a
tion of the basilar membrane excites the hair endings.
Figure 52–8 shows the mechanism by which vibra-
ing with their bases.
This in turn excites the auditory nerve fibers synaps-
hairs in one direction depolarizes the hair cells, and
which are discussed in Chapter 55. Bending of the
and cristae ampullaris of the vestibular apparatus,
lies above the stereocilia in the scala media. These hair
tectorial membrane,
minute hairs, or
Note in Figure 52–7 that
cochlear nerve is shown in Figure 52–2.
system at the level of the upper medulla. The relation
cochlear nerve
(center) of the cochlea. The spiral ganglion neuronal
The nerve fibers stimulated by the hair cells lead to
endings terminate on the inner hair cells, which
nerve endings. Between 90 and 95 per cent of these
of only about 8 micrometers. The bases and sides of
cells,
ter, and three or four rows of
hair cells,
of the basilar fibers and basilar membrane. The actual
brane. Note that the organ of Corti lies on the surface
52–7, is the receptor organ that generates nerve
The organ of Corti, shown in Figures 52–2, 52–3, and
from the organ of Corti lying on the basilar membrane,
“place” of maximum stimulation of the nerve fibers
The principal method by which sound frequencies
helicotrema, where the scala vestibuli opens into the
that for frequencies less than 200 cycles per second is
second occurs near the base of the cochlea, whereas
maximum amplitude for sound at 8000 cycles per
Chapter 52
The Sense of Hearing
655
all the way at the tip of the basilar membrane near the
scala tympani.
are discriminated from one another is based on the
as explained in the next section.
Function of the Organ of Corti
impulses in response to vibration of the basilar mem-
sensory receptors in the organ of Corti are two spe-
cialized types of nerve cells called hair cells—a single
row of internal (or “inner”)
numbering about
3500 and measuring about 12 micrometers in diame-
external (or “outer”) hair
numbering about 12,000 and having diameters
the hair cells synapse with a network of cochlea
emphasizes their special importance for the detection
of sound.
the spiral ganglion of Corti, which lies in the modiolus
cells send axons—a total of about 30,000—into the
and then into the central nervous
of the organ of Corti to the spiral ganglion and to the
Excitation of the Hair Cells.
stereocilia, project upward from the
hair cells and either touch or are embedded in the
surface gel coating of the
which
cells are similar to the hair cells found in the macula
bending in the opposite direction hyperpolarizes them.
retic-
ular lamina, supported by triangular rods of Corti,
lamina move as a rigid unit.
Upward movement of the basilar fiber rocks the
reticular lamina upward and
toward the modi-
hairs on the hair cells to shear back and forth against
Even though there are three to four times as many
the auditory nerve fibers are stimulated by the inner
the outer cells are damaged while the inner cells
outer hair cells in some way control the sensitivity
Tectorial membrane
Basilar fiber
Outer hair cells
Inner hair cells
Spiral ganglion
Cochlear nerve
membrane pressing against the projecting hairs.
Organ of Corti, showing especially the hair cells and the tectorial
Figure 52–7
Hairs
Tectorial membrane
Basilar fiber
Reticular lamina
Rods of Corti
Modiolus
projecting into the gel coating of the tectorial membrane.
Stimulation of the hair cells by to-and-fro movement of the hairs
Figure 52–8

at more rapid rates.
increases, so that the hair cells excite the nerve endings
First, as the sound becomes louder, the amplitude of
least three ways.
Determination of Loudness
tion of the lower-frequency sounds.
mally detected, does not totally eliminate discrimina-
membrane where all lower-frequency sounds are nor-
apical half of the cochlea, which destroys the basilar
cies of the volleys. In fact, destruction of the entire
nuclei of the brain. It is further suggested that the
nized at the same frequencies, and these volleys are
second, can cause volleys of nerve impulses synchro-
frequency sounds, from 20 to 1500 to 2000 cycles per
That is, low-
frequency principle.
range from 200 down to 20. It is postulated that these
200 cycles per second. Therefore, it has been difficult
Yet, referring again to Figure 52–6, one can see that
frequency.
membrane that are most stimulated. This is called
frequencies. Therefore, the
cochlea to the cerebral cortex. Recording of signals in
fibers in the cochlear pathway, all the way from the
Furthermore, there is spatial organization of the nerve
mediate distances between the two extremes.
frequency sounds activate the membrane at inter-
brane near the base of the cochlea. Intermediate-
and high-frequency sounds activate the basilar mem-
that low-frequency sounds cause maximal activation
From earlier discussions in this chapter, it is apparent
Principle
extra amount, thereby increasing its ability to respond
endolymph. It is believed that this high electrical poten-
the hair cells. Furthermore, the hair cells have a nega-
media, whereas perilymph bathes the lower bodies of
The importance of the endocochlear potential is that
into the scala media by the stria vascularis.
endocochlear potential,
This is called the
positivity inside the scala media and negativity outside.
all the time between endolymph and perilymph, with
of sodium, which is exactly opposite to the contents of
wall of the scala media. Endolymph contains a high
stria vascularis,
brospinal fluid. Conversely, the endolymph that fills
The scala vestibuli and scala tympani communicate
The scala media is filled with a
endocochlear potential:
electrical potentials generated by the hair cells, we need
To explain even more fully the
sible that the transmitter substance is glutamate, but
cells at these synapses during depolarization. It is pos-
with the bases of the hair cells. It is believed that a
alternating hair cell receptor potential. This, in turn,
direction they hyperpolarize, thereby generating an
vestibuli, the hair cells depolarize, and in the opposite
Thus, when the basilar fibers bend toward the scala
ization of the hair cell membrane.
media fluid into the stereocilia, which causes depolar-
channels, allowing rapid movement of positively
the surface of the hair cell. This causes a mechanical
cilia are bent in the direction of the longer ones, the
adjacent longer stereocilia. Therefore, whenever the
the modiolus, and the tops of the shorter stereocilia are
100 stereocilia on its apical border. These become pro-
has a rigid protein framework. Each hair cell has about
The stereocilia (the “hairs” protruding from the
Hair Cell Receptor Potentials and Excitation of Auditory Nerve
pitches, activated through the outer hair cells.
for control of the ear’s sensitivity to different sound
These effects suggest a retrograde nervous mechanism
cells and possibly also change their degree of stiffness.
vicinity of the outer hair cells. Stimulating these nerve
In support of this concept, a large number of retro-
phenomenon called “tuning” of the receptor system.
The Nervous System: B. The Special Senses
656
Unit X
grade nerve fibers pass from the brain stem to the
fibers can actually cause shortening of the outer hair
Fibers.
ends of the hair cells) are stiff structures because each
gressively longer on the side of the hair cell away from
attached by thin filaments to the back sides of their
tips of the smaller stereocilia are tugged outward from
transduction that opens 200 to 300 cation-conducting
charged potassium ions from the surrounding scala
stimulates the cochlear nerve endings that synapse
rapidly acting neurotransmitter is released by the hair
this is not certain.
Endocochlear Potential.
to explain another electrical phenomenon called the
fluid called endolymph, in contradistinction to the peri-
lymph present in the scala vestibuli and scala tympani.
directly with the subarachnoid space around the brain,
so that the perilymph is almost identical with cere-
the scala media is an entirely different fluid secreted by
the
a highly vascular area on the outer
concentration of potassium and a low concentration
perilymph.
An electrical potential of about +80 millivolts exists
and it is gen-
erated by continual secretion of positive potassium ions
the tops of the hair cells project through the reticular
lamina and are bathed by the endolymph of the scala
tive intracellular potential of –70 millivolts with respect
to the perilymph but –150 millivolts with respect to the
endolymph at their upper surfaces where the hairs
project through the reticular lamina and into the
tial at the tips of the stereocilia sensitizes the cell an
to the slightest sound.
Determination of Sound Frequency—
The “Place”
of the basilar membrane near the apex of the cochlea,
the auditory tracts of the brain stem and in the audi-
tory receptive fields of the cerebral cortex shows that
specific brain neurons are activated by specific sound
major method used by
the nervous system to detect different sound frequen-
cies is to determine the positions along the basilar
the place principle for the determination of sound
the distal end of the basilar membrane at the heli-
cotrema is stimulated by all sound frequencies below
to understand from the place principle how one can
differentiate between low sound frequencies in the
low frequencies are discriminated mainly by the so-
called volley or
transmitted by the cochlear nerve into the cochlear
cochlear nuclei can distinguish the different frequen-
Loudness is determined by the auditory system in at
vibration of the basilar membrane and hair cells also
body, (2) in the commissure between the two nuclei of
occurs between the two pathways: (1) in the trapezoid
In at least three places in the brain stem, crossing over
derance of transmission in the contralateral pathway.
pathways of both sides of the brain, with a prepon-
Several important points should be noted. First,
gyrus of the temporal lobe.
where all the fibers do synapse. Finally, the
nucleus,
there, the pathway passes to the
all or almost all the auditory fibers synapse. From
nucleus and travel on to the inferior colliculus, where
nucleus of the lateral lemniscus,
lateral lemniscus.
on the same side. From the superior olivary nucleus,
superior olivary nucleus.
all the fibers synapse, and second-order neurons pass
located in the upper part of the medulla. At this point,
ventral cochlear nuclei
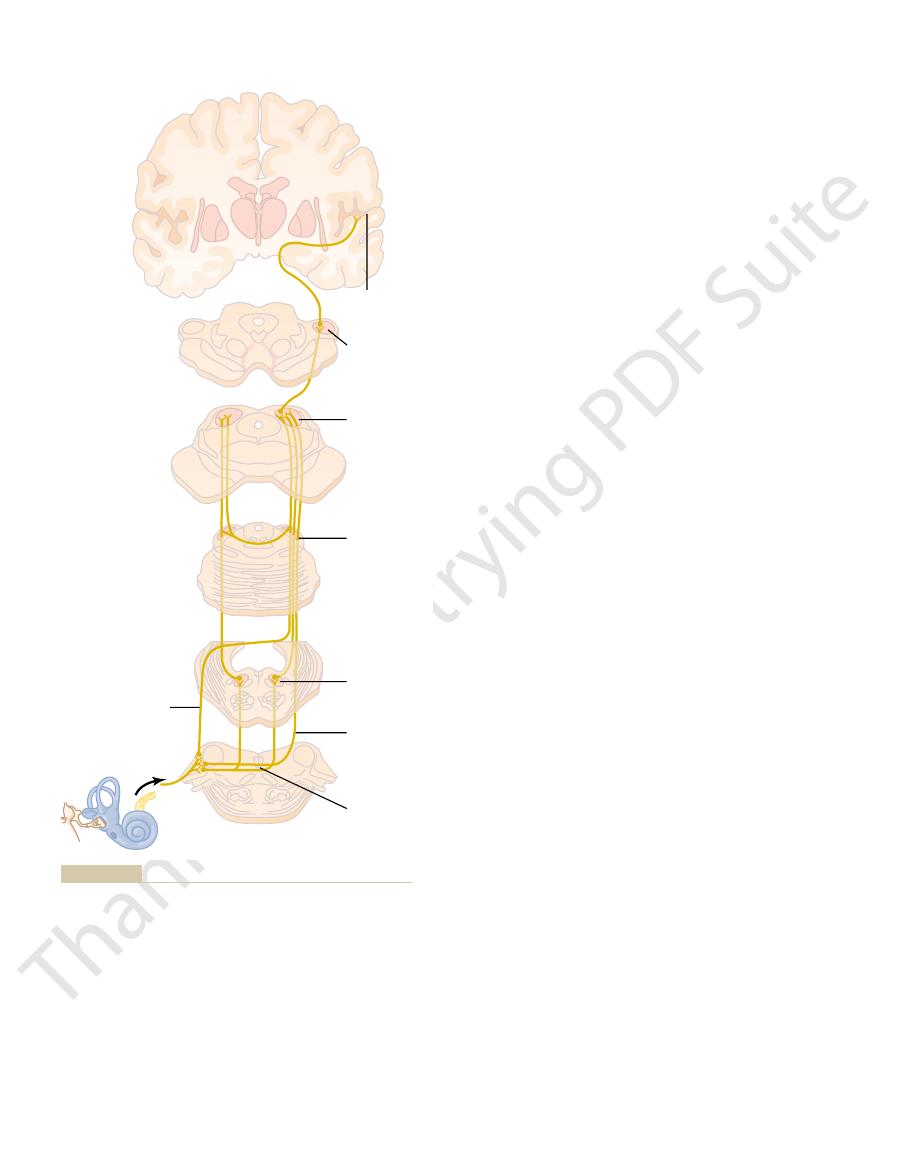
Figure 52–10 shows the major auditory pathways. It
Central Auditory Mechanisms
the chapter.
50 to 8000 cycles per second or less, as discussed later in
In old age, this frequency range is usually shortened to
the complete range of 20 to 20,000 cycles be achieved.
to 5000 cycles per second; only with intense sounds can
sound pressure level, the sound range is 500
extent on loudness. If the loudness is 60 decibels below
52–9, we see that the sound range depends to a great
cycles per second. However, referring again to Figure
The frequencies of sound that
as this.
timeter. Conversely, a 100-cycle-per-second sound can
This figure demonstrates that a 3000-cycle-per-second
different frequencies can barely be heard by the ear.
Figure
sound intensity.
change
intensity range for communication, the ears can barely
express changes in loudness is that, in the usual sound
sound energy of 1.26 times.
and 0.1 bel is called 1
logarithm of their actual intensities. A 10-fold increase
intensities that the ear can detect and discriminate,
compression of the intensity scale.
system. This allows a person to interpret differences in
Thus, the scale of intensity is greatly “compressed” by
in sound level as approximately a 10,000-fold change.
membrane. Yet the ear interprets this much difference
approximately 1 trillion
possible noise, representing an
another way, the ear can discriminate differences in
root of the actual sound intensity. To express this
intensity. In the case of sound, the interpreted sensa-
pointed out in Chapter 46, a person interprets changes
Detection of Changes in Loudness—The Power Law.
reaches high intensity, and stimulation of these cells
Third, the outer hair cells do not become stimulated
fibers rather than through only a few.
impulses—that is, transmission through many nerve
become stimulated, thus causing
Second, as the amplitude of vibration increases, it
Chapter 52
The Sense of Hearing
657
causes more and more of the hair cells on the fringes
of the resonating portion of the basilar membrane to
spatial summation of
significantly until vibration of the basilar membrane
presumably apprises the nervous system that the
sound is loud.
As
in intensity of sensory stimuli approximately in pro-
portion to an inverse power function of the actual
tion changes approximately in proportion to the cube
sound intensity from the softest whisper to the loudest
times increase in sound energy or 1 million times
increase in amplitude of movement of the basilar
the sound perception mechanisms of the auditory
sound intensities over an extremely wide range—a far
wider range than would be possible were it not for
Decibel Unit.
Because of the extreme changes in sound
sound intensities are usually expressed in terms of the
in sound energy is called 1 bel,
decibel. One decibel represents an actual increase in
Another reason for using the decibel system to
distinguish an approximately 1-decibel
in
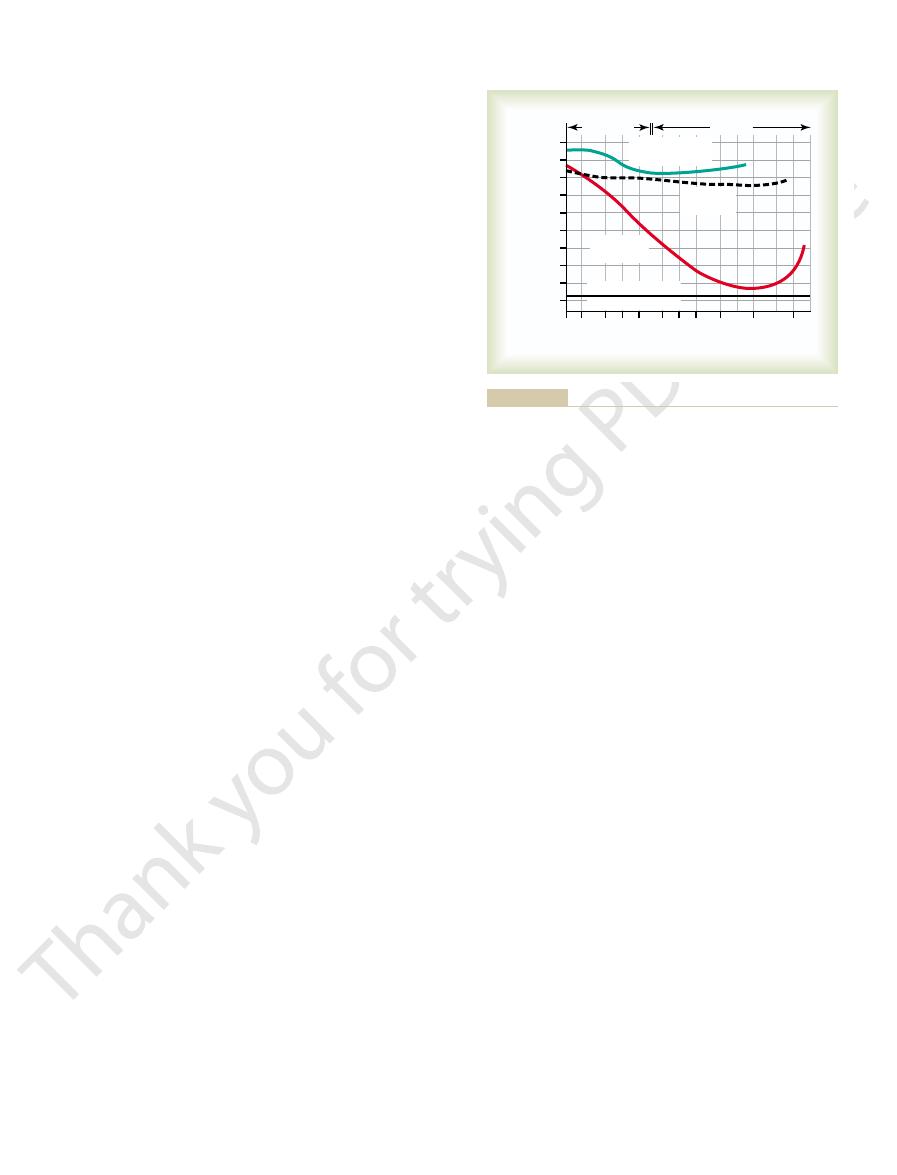
Threshold for Hearing Sound at Different Frequencies.
52–9 shows the pressure thresholds at which sounds of
sound can be heard even when its intensity is as low as
70 decibels below 1 dyne/cm
2
sound pressure level,
which is one ten-millionth microwatt per square cen-
be detected only if its intensity is 10,000 times as great
Frequency Range of Hearing.
a young person can hear are between 20 and 20,000
1 dyne/cm
2
Auditory Nervous Pathways
shows that nerve fibers from the spiral ganglion of
Corti enter the dorsal and
mainly to the opposite side of the brain stem to ter-
minate in the
A few second-
order fibers also pass to the superior olivary nucleus
the auditory pathway passes upward through the
Some of the fibers terminate in the
but many bypass this
medial geniculate
pathway proceeds by way of the auditory radiation to
the auditory cortex, located mainly in the superior
signals from both ears are transmitted through the
5 10 20
100
500 2000 10,000
Vibration
Sound
Pressure in db
(0 decibel = 1 dyne/cm
2
)
Frequency
1 2
– 80
–60
–40
–20
0
20
40
60
80
100
Pricking
(in middle ear)
Tactual
threshold
Threshold
of hearing
Reference
pressure =
-
73.8
each sound frequency.
tion (pricking and tactual threshold) to the sound energy level at
Relation of the threshold of hearing and of somesthetic percep-
Figure 52–9
sounds are located posteriorly. This is not true for all
orly, as shown in Figure 52–11, and the high-frequency
In most, the low-frequency sounds are located anteri-
frequency sounds excite neurons at the opposite end.
excite neurons at one end of the map, whereas low-
areas. In each of these maps, high-frequency sounds
Sound Frequency Perception in the Primary Auditory Cortex.
body.
by projections from the medial geniculate body,
). The primary auditory cortex is directly excited
52–11: the
Two separate subdivisions are shown in Figure
, and even onto the
lateral side of the temporal lobe,
cortex is shown in Figure 52–11, which demonstrates
The projection area of auditory signals to the cerebral
in Hearing
Function of the Cerebral Cortex
direction from which sound comes.
about this later, especially in relation to perception of
as low as the cochlear nuclei. We will have more to say
the brain; instead, information from the sound signals
this synchronization is mainly lost. These findings
per second.Above the level of the inferior colliculi, even
quency, except at sound frequencies below 200 cycles
In the auditory tracts of the brain stem, the firing is
not necessarily occur with every wave.
often synchronized with the sound waves, but they do
4000 cycles per second, the auditory nerve impulses are
ness of the sound. At sound frequencies up to 2000 to
second, the rate being determined mainly by the loud-
Firing Rates at Different Levels of the Auditory Pathways.
tion areas.
the auditory cortex, and
one precise pattern
two patterns
to the cortex. In fact, there are
Third, a high degree of spatial orientation is main-
taneously in the event of a sudden noise.
response to loud sounds. Other collaterals go to the
This system projects diffusely
Second, many collateral fibers from the auditory
the lateral lemnisci, and (3) in the commissure con-
The Nervous System: B. The Special Senses
658
Unit X
necting the two inferior colliculi.
tracts pass directly into the reticular activating system
of the brain stem.
upward in the brain stem and downward into the
spinal cord and activates the entire nervous system in
vermis of the cerebellum, which is also activated instan-
tained in the fiber tracts from the cochlea all the way
three spatial patterns for
termination of the different sound frequencies in the
cochlear nuclei,
in the inferior colliculi,
for discrete sound frequencies in
at least five other less precise
patterns in the auditory cortex and auditory associa-
Single
nerve fibers entering the cochlear nuclei from the audi-
tory nerve can fire at rates up to at least 1000 per
usually no longer synchronized with the sound fre-
demonstrate that the sound signals are not transmitted
unchanged directly from the ear to the higher levels of
begins to be dissected from the impulse traffic at levels
that the auditory cortex lies principally on the
supratemporal plane of the superior temporal gyrus but
also extends onto the
over much of the insular cortex
lateral portion of the parietal operculum.
primary auditory cortex and the auditory
association cortex (also called the secondary auditory
cortex
whereas the auditory association areas are excited
secondarily by impulses from the primary auditory
cortex as well as by some projections from thalamic
association areas adjacent to the medial geniculate
At least six tonotopic maps have been found in the
primary auditory cortex and auditory association
Primary
auditry
cortex
Medial
geniculate
nucleus
Inferior
colliculus
Nucleus of
the lateral
lemniscus
Superior
olivary
nuclei
Intermediate
acoustic
site
Medulla
Trapezoid
body
Midbrain
Midbrain
Pons
Pons
Dorsal
acoustic
stria
Cochlear
nuclei
N. VIlI
3rd ed. New York: Oxford University Press, 1981.)
system. In Neurological Anatomy in Relation to Clinical Medicine,
Auditory nervous pathways. (Modified from Brodal A: The auditory
Figure 52–10

Chapter 57.
repeat them. These functions of the auditory associa-
tory association cortex, often make it impossible for a
which is called Wernicke’s area and is part of the audi-
the posterior portion of the superior temporal gyrus,
of the sound heard. For instance, lesions in
However, he or she is often unable to interpret the
person’s ability to hear and differentiate sound tones
localize the source of a sound, because comparative
pathway. However, it does affect one’s ability to
reduces hearing in the opposite ear; it does not cause
for hearing. Destruction of one side only slightly
the human being greatly reduces one’s sensitivity
patterns.
Therefore, the auditory cortex is especially important
more, the animal cannot relearn this type of response.
ability when the auditory cortex is destroyed; further-
lowing the other in a particular pattern, loses this
ognize a combination or sequence of tones, one fol-
For instance, an animal that has been trained to rec-
even abolish the animal’s ability to discriminate dif-
However, it does greatly reduce or sometimes
sounds or reacting in a crude manner to sounds.
Discrimination of Sound “Patterns” by the Auditory Cortex.
sory area II, which could provide an easy opportunity
areas of the cortex. Indeed, the parietal portion of the
believed that these neurons “associate” different
only to specific sound frequencies in the ear. It is
sensations.
somesthetic images, visual images, and other types of
adjacent pathways. The same effect has been demon-
on both sides of this primary frequency; this is
cochlea at one frequency inhibits sound frequencies
ting information in nerves. That is, stimulation of the
nomenon of lateral inhibition, which is discussed
“sharpen” the frequency response. It is believed that
where along the pathway, processing mechanisms
cies rather than to a broad range. Therefore, some-
reached the cerebral cortex, most sound-responsive
tation is found. Yet, by the time the excitation has
cochlear nuclei, this same breadth of sound represen-
stimulated by sounds of all frequencies, and in the
relay nuclei. Referring back to Figure 52–6
The frequency range to which each individual
pure frequency sounds.
or perhaps special modulations, such as noise versus
special qualities, such as the sudden onset of sounds,
sound comes. Other auditory cortex areas detect
the psychic sensation of sound pitches. Another map
sounds. For instance, one of the large maps in the
maps? The answer, presumably, is that each of the sep-
the maps. The question one must ask is, Why does the
Chapter 52
The Sense of Hearing
659
auditory cortex have so many different tonotopic
arate areas dissects out some specific feature of the
primary auditory cortex almost certainly discriminates
the sound frequencies themselves and gives the person
is probably used to detect the direction from which the
neuron in the auditory cortex responds is much
narrower than that in the cochlear and brain stem
B, note that
the basilar membrane near the base of the cochlea is
neurons respond to only a narrow range of frequen-
this sharpening effect is caused mainly by the phe-
in Chapter 46 in relation to mechanisms for transmit-
caused by collateral fibers angling off the primary
signal pathway and exerting inhibitory influences on
strated to be important in sharpening patterns of
Many of the neurons in the auditory cortex, espe-
cially in the auditory association cortex, do not respond
sound frequencies with one another or associate sound
information with information from other sensory
auditory association cortex partly overlaps somatosen-
for the association of auditory information with
somatosensory information.
Complete bilateral removal of the auditory cortex
does not prevent a cat or monkey from detecting
ferent sound pitches and especially patterns of sound.
in the discrimination of tonal and sequential sound
Destruction of both primary auditory cortices in
deafness in the ear because of many crossover con-
nections from side to side in the auditory neural
signals in both cortices are required for the localiza-
tion function.
Lesions that affect the auditory association areas
but not the primary auditory cortex do not decrease a
or even to interpret at least simple patterns of sound.
meaning
person to interpret the meanings of words even though
he or she hears them perfectly well and can even
tion areas and their relation to the overall intellectual
functions of the brain are discussed in more detail in
Low
frequency
High
frequency
Primary
Primary
Association
Association
Auditory cortex.
Figure 52–11

abilities, the “audiometer” is used. Simply an earphone
To determine the nature of hearing dis-
Audiometer.
applied to the skull over the ear.
ankylosed (“frozen” in place by fibrosis or calcification),
However, if the cochlea and nerve are still intact but the
destroyed, the person becomes permanently deaf.
deafness.”
itself to the cochlea, which is usually called “conduction
deafness,” and (2) that caused by impairment of the
the auditory nerve, which is usually classified as “nerve
Deafness is usually divided into two types: (1) that
Types of Deafness
Hearing Abnormalities
ties. This is readily demonstrated when one listens to a
bels. One can readily understand how this could allow
Corti, reducing their sound sensitivities 15 to 20 deci-
These retrograde fibers are inhibitory. Indeed, direct
the cochlea in the ear itself. The final pathway is mainly
Central Nervous System to Lower
Centrifugal Signals from the
“quality” of sound tones at the level of the superior
“quality” of sound direction is separated from the
different levels of neuronal activity. In this case, the
This mechanism for detection of sound direction
stimulated neurons. It is believed that all these signals
then transmitted to the auditory cortex, where sound
on opposite sides. This spatial orientation of signals is
develops in the medial superior olivary nucleus, with
lags. Thus, a spatial pattern of neuronal stimulation
lag; those in between respond to intermediate time
nucleus respond maximally to a short time lag, while
from the two ears. The neurons near one border of the
the left ear impinges on the left dendrite. The intensity
ear impinges on the right dendrite, and the signal from
other to the left. The acoustical signal from the right
major dendrites, one projecting to the right and the
This nucleus
acoustical signals entering the two ears.
however, has a
medial superior olivary nucleus,
The
the sound is coming, presumably by simply comparing
The lateral nucleus
lateral superior olivary nucleus.
tions: (1) the
The superior olivary nucleus is divided into two sec-
believed to be the following.
for interpretation of the signals. The mechanism is
in the brain stem, even though the neural pathways all
which sound comes. Yet the neural analyses for this
whether in human beings or in lower mammals, causes
ferent directions.
tion from which the sound comes. It does this by
of the sound entering the ear, depending on the direc-
two ears. The shape of the pinna changes the
behind the person or from above or below. This dis-
The two aforementioned mechanisms cannot tell
enter the brain ahead of those from the left ear.
than the left ear is, the sound signals from the right ear
instant, whereas if the right ear is closer to the sound
sound, the sound reaches both ears at exactly the same
exact interval of time between two acoustical signals.
The time lag mechanism discriminates direction much
head is a greater sound barrier at these frequencies.
below 3000 cycles per second, and the second mecha-
The first mechanism functions best at frequencies
between the intensities of the sounds in the two ears.
its entry into the opposite ear, and (2) the difference
which sound comes by two principal means: (1) the
Determination of the Direction from
The Nervous System: B. The Special Senses
660
Unit X
Which Sound Comes
A person determines the horizontal direction from
time lag between the entry of sound into one ear and
nism operates best at higher frequencies because the
more exactly than the intensity mechanism because it
does not depend on extraneous factors but only on the
If a person is looking straight toward the source of the
whether the sound is emanating from in front of or
crimination is achieved mainly by the pinnae of the
quality
emphasizing specific sound frequencies from the dif-
Neural Mechanisms for Detecting Sound Direction.
Destruc-
tion of the auditory cortex on both sides of the brain,
loss of almost all ability to detect the direction from
detection process begin in the superior olivary nuclei
the way from these nuclei to the cortex are required
medial superior olivary nucleus and (2)
the
is concerned with detecting the direction from which
the difference in intensities of the sound reaching the
two ears and sending an appropriate signal to the audi-
tory cortex to estimate the direction.
specific mechanism for detecting the time lag between
contains large numbers of neurons that have two
of excitation of each neuron is highly sensitive to a
specific time lag between the two acoustical signals
those near the opposite border respond to a long time
sound from directly in front of the head stimulating
one set of olivary neurons maximally and sounds from
different side angles stimulating other sets of neurons
direction is determined by the locus of the maximally
for determining sound direction are transmitted
through a different pathway and excite a different
locus in the cerebral cortex from the transmission
pathway and termination locus for tonal patterns of
sound.
indicates again how specific information in sensory
signals is dissected out as the signals pass through
olivary nuclei.
Auditory Centers
Retrograde pathways have been demonstrated at each
level of the auditory nervous system from the cortex to
from the superior olivary nucleus to the sound-receptor
hair cells in the organ of Corti.
stimulation of discrete points in the olivary nucleus has
been shown to inhibit specific areas of the organ of
a person to direct his or her attention to sounds of par-
ticular qualities while rejecting sounds of other quali-
single instrument in a symphony orchestra.
caused by impairment of the cochlea or impairment of
physical structures of the ear that conduct sound
If either the cochlea or the auditory nerve is
tympanum-ossicular system has been destroyed or
sound waves can still be conducted into the cochlea by
means of bone conduction from a sound generator
of complex sound: a way forward? Trends Neurosci 27:181,
Griffiths TD, Warren JD, Scott SK, et al: Cortical processing
motor. Annu Rev Physiol 66:521, 2004.
Gillespie PG, Cyr JL: Myosin-1c, the hair cell’s adaptation
Mammalian Ear. New York: Oxford University Press,
Geisler CD: From Sound to Synapse: Physiology of the
cells. Nat Rev Genet 5:489, 2004.
Frolenkov GI, Belyantseva IA, Friedman TB, Griffith AJ:
Opin Neurobiol 13:446, 2003.
Fettiplace R, Ricci AJ:Adaptation in auditory hair cells. Curr
Curr Biol 13:R767, 2003.
channels.
Eatock RA: Auditory physiology: listening with K
ing. New York: Plenum Press, 1998.
Brugge J: Central Auditory Processing and Neural Model-
sound from the incus to the oval window.
a minute Teflon or metal prosthesis that transmits the
this case, the person becomes totally deaf for ossicular
bone overgrowth to the edges of the oval window. In
the faceplate of the stapes becomes “ankylosed” by
frequencies. In some instances of conduction deafness,
greatly depressed at all frequencies, but more so at low
normal, but conduction through the ossicular system is
deafness.” In this case, bone conduction is essentially
gram from a person with “middle ear air conduction
brane to the oval window. Figure 52–13 shows an audio-
In either case, the sound waves cannot be transmitted
as streptomycin, kanamycin, and chloramphenicol.
Corti—in particular, sensitivity to some antibiotics such
aging to the organ of Corti, and (2) deafness for all
low-frequency sounds are usually louder and more dam-
sounds (a rock band or a jet airplane engine), because
follows: (1) deafness for low-frequency sounds caused
all older people.
This type of deafness occurs to some extent in almost
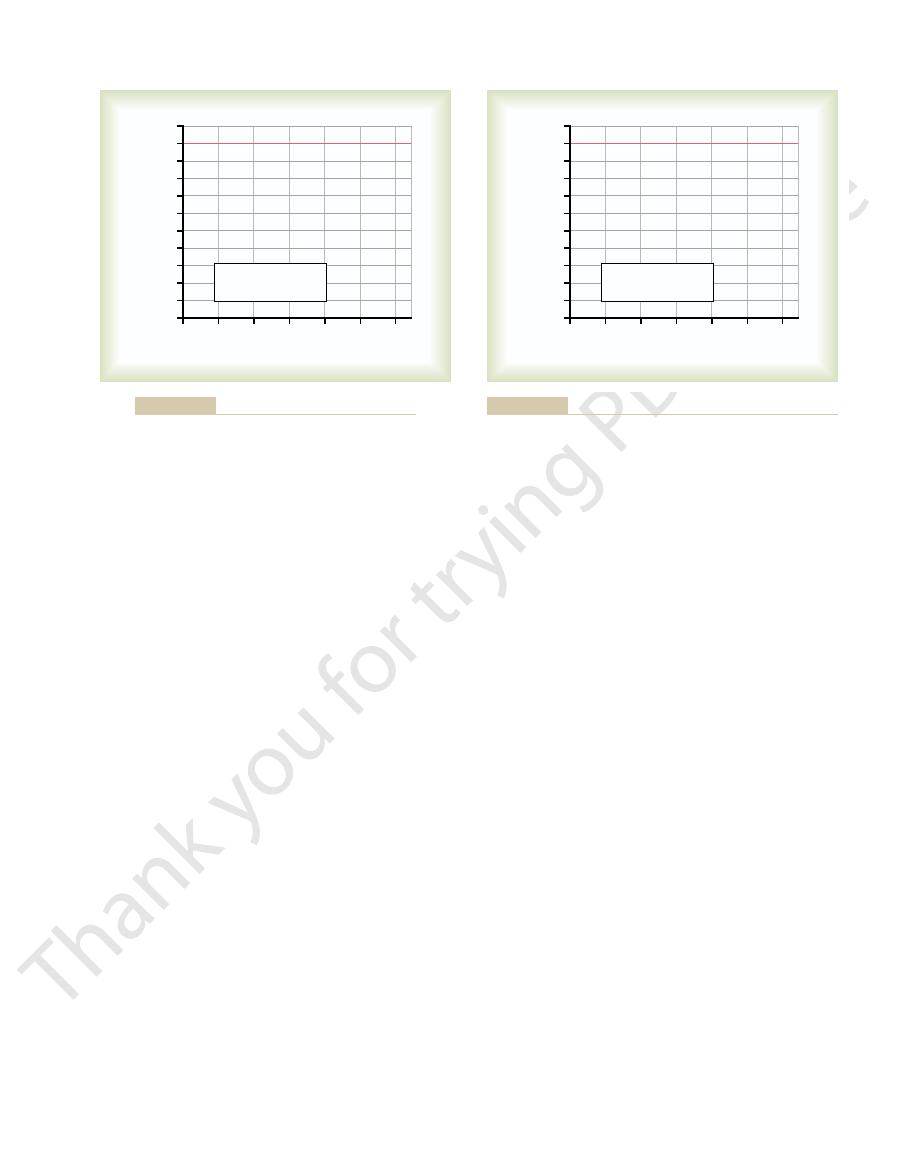
ness is mainly for high-frequency sound. Such deafness
ness is shown in Figure 52–12. In this figure, the deaf-
conduction. An audiogram depicting partial nerve deaf-
nerve, or the central nervous system circuits from the
which includes damage to the cochlea, the auditory
In nerve deafness—
by the ear, is equipped with a mechanical vibrator for
spectrum. The audiometer, in addition to being
plotted, as shown in Figures 52–12 and 52–13, depicting
these frequencies. Then the so-called
spectrum, and the hearing loss is determined for each of
In performing a hearing test using an audiometer, one
ular frequency.
bels above normal before it can be heard, the person is
zero level. If the loudness must be increased to 30 deci-
that can barely be heard by the normal ear. A calibrated
intensity-level sound at each frequency is the loudness
quencies, the instrument is calibrated so that zero-
Chapter 52
The Sense of Hearing
661
connected to an electronic oscillator capable of emitting
pure tones ranging from low frequencies to high fre-
volume control can increase the loudness above the
said to have a hearing loss of 30 decibels at that partic-
tests about 8 to 10 frequencies covering the auditory
audiogram is
hearing loss at each of the frequencies in the auditory
equipped with an earphone for testing air conduction
testing bone conduction from the mastoid process of the
skull into the cochlea.
Audiogram in Nerve Deafness.
ear—the person has decreased or total loss of ability to
hear sound as tested by both air conduction and bone
could be caused by damage to the base of the cochlea.
Other patterns of nerve deafness frequently occur as
by excessive and prolonged exposure to very loud
frequencies caused by drug sensitivity of the organ of
Audiogram for Middle Ear Conduction Deafness.
A
common type of deafness is caused by fibrosis in the
middle ear following repeated infection or by fibrosis
that occurs in the hereditary disease called otosclerosis.
easily through the ossicles from the tympanic mem-
conduction but can regain almost normal hearing by the
surgical removal of the stapes and its replacement with
References
+
Genetic insights into the morphogenesis of inner ear hair
1998.
2004.
1
250
500
1000 2000 4000 8000
Loss in decibels
Frequency
100
90
80
70
60
50
40
30
20
-
10
10
Normal
X
Air conduction
X
*
Bone conduction
*
X
*
X
*
X
*
X
*
X
X
*
Figure 52–12
Audiogram of the old-age type of nerve deafness.
125
250
500
1000 2000 4000 8000
Loss in decibels
Frequency
100
90
80
70
60
50
40
30
20
-
10
10
Normal
X
Air conduction
X
*
Bone conduction
*
X
*
X
*
X
*
X
*
X
X
*
sclerosis.
Audiogram of air conduction deafness resulting from middle ear
Figure 52–13

primary auditory cortex. Nat Rev Neurosci 5:279, 2004.
Weinberger NM: Specific long-term memory traces in
999:506, 2003.
ing on the auditory cortex in children. Ann N Y Acad Sci
Trainor LJ, Shahin A, Roberts LE: Effects of musical train-
Physiol Rev 82:601, 2002.
hearing loss, restoration of function, and during learning.
Syka J: Plastic changes in the central auditory system after
plasticity in the auditory system. Nat Rev Neurosci 4:783,
Suga N, Ma X: Multiparametric corticofugal modulation and
Opin Neurobiol 13:167, 2003.
Semple MN, Scott BH: Cortical mechanisms in hearing. Curr
100, 2003.
organization of speech perception. Trends Neurosci 26:
Scott SK, Johnsrude IS: The neuroanatomical and functional
Rev Neurosci 23:501, 2000.
of frequency integration in primary auditory cortex. Annu
Schreiner CE, Read HL, Sutter ML: Modular organization
cell boogie rules. Curr Opin Neurobiol 13:459, 2003.
Santos-Sacchi J: New tunes from Corti’s organ: the outer hair
cochlea. Physiol Rev 81:1305, 2001.
Robles L, Ruggero MA: Mechanics of the mammalian
of auditory cortex. Curr Opin Neurobiol 12:433, 2002.
Read HL, Winer JA, Schreiner CE: Functional architecture
Science 295:1025, 2002.
Rauschecker JP, Shannon RV: Sending sound to the brain.
Opin Neurol 17:9, 2004.
Minor LB, Schessel DA, Carey JP: Meniere’s disease. Curr
the noise? Curr Biol 14:R231, 2004.
McDermott BM Jr, Lopez-Schier H: Inner ear: Ca
tude-modulated sounds. Physiol Rev 84:541, 2004.
Joris PX, Schreiner CE, Rees A: Neural processing of ampli-
ization. Nat Rev Neurosci 4:540, 2003.
Grothe B: New roles for synaptic inhibition in sound local-
The Nervous System: B. The Special Senses
662
Unit X
2+
you feel
2003.