
in the cones, it is one of three “color”
In the case of the rods, this is
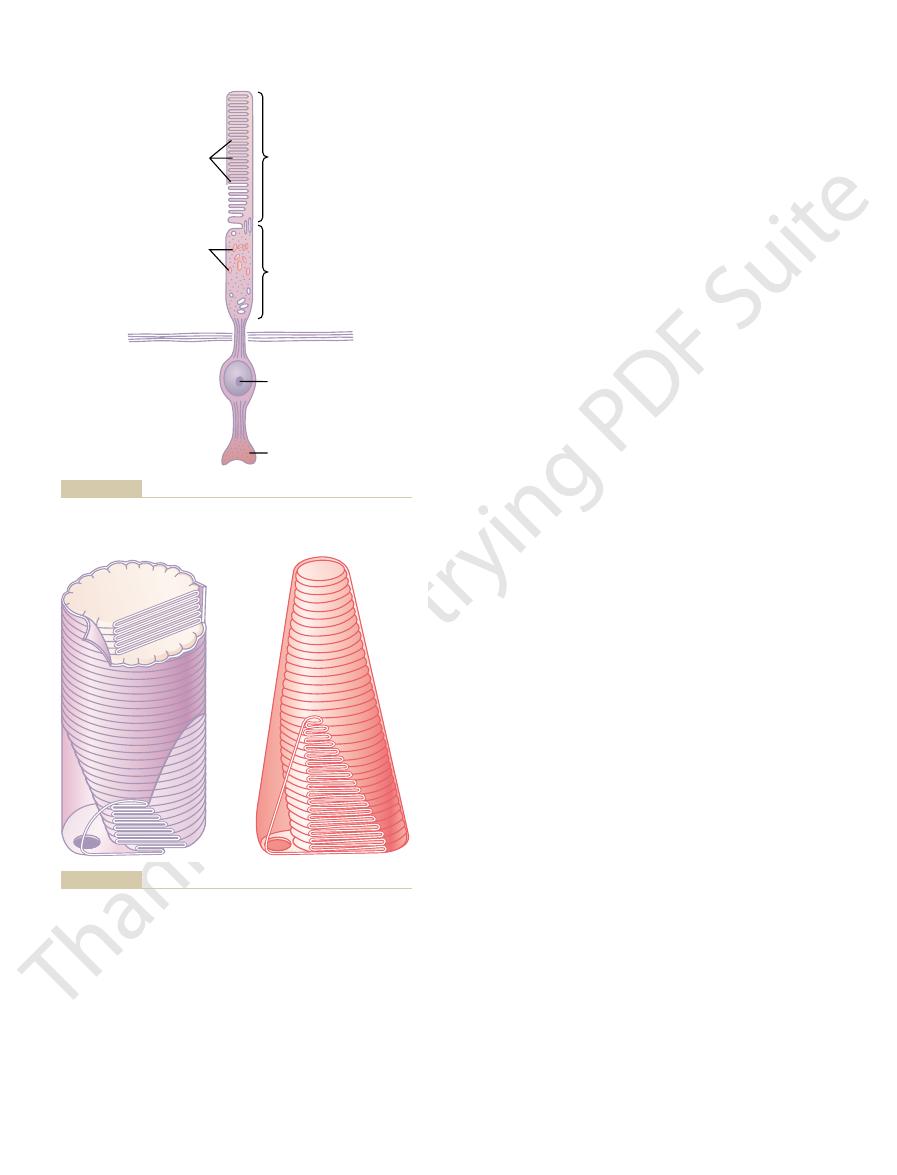
The light-sensitive photochemical is found in the outer segment.
synaptic body.
nucleus,
rod or a cone: (1) the
To the right in Figure 50–3 are labeled the major functional segments of either a
rods, and the cones are slender and have a diameter of only 1.5 micrometers.
8 micrometers in diameter; in the central part of the retina, in the fovea, there are
the retina, the rods are 2 to 5 micrometers in diameter, whereas the cones are 5 to
longer than the cones, but this is not always the case. In the peripheral portions of
outer segment of the cone is conical in shape. In general, the rods are narrower and
ponents of a photoreceptor (either a rod or a cone). As shown in Figure 50–4, the
Figure 50–3 is a diagrammatic representation of the essential com-
of the cones. This allows light to pass unimpeded to the cones.
in the foveal region, the blood vessels, ganglion cells, inner nuclear layer of cells,
tradistinction to the much fatter cones located more peripherally in the retina. Also,
image. That is, the foveal cones have especially long and slender bodies, in con-
only 0.3 millimeter in diameter, is composed almost entirely of cones;
central fovea,
than 1 square millimeter; it is especially capable of acute and detailed vision. The
in the center of the retina, shown in Figure 50–2, occupying a total area a little more
fovea
The
are pulled aside to decrease this loss of acuity.
in the central foveal region of the retina, as discussed subsequently, the inside layers
acuity is decreased by this passage through such nonhomogeneous tissue. However,
of the retina. This distance is a thickness of several hundred micrometers; visual
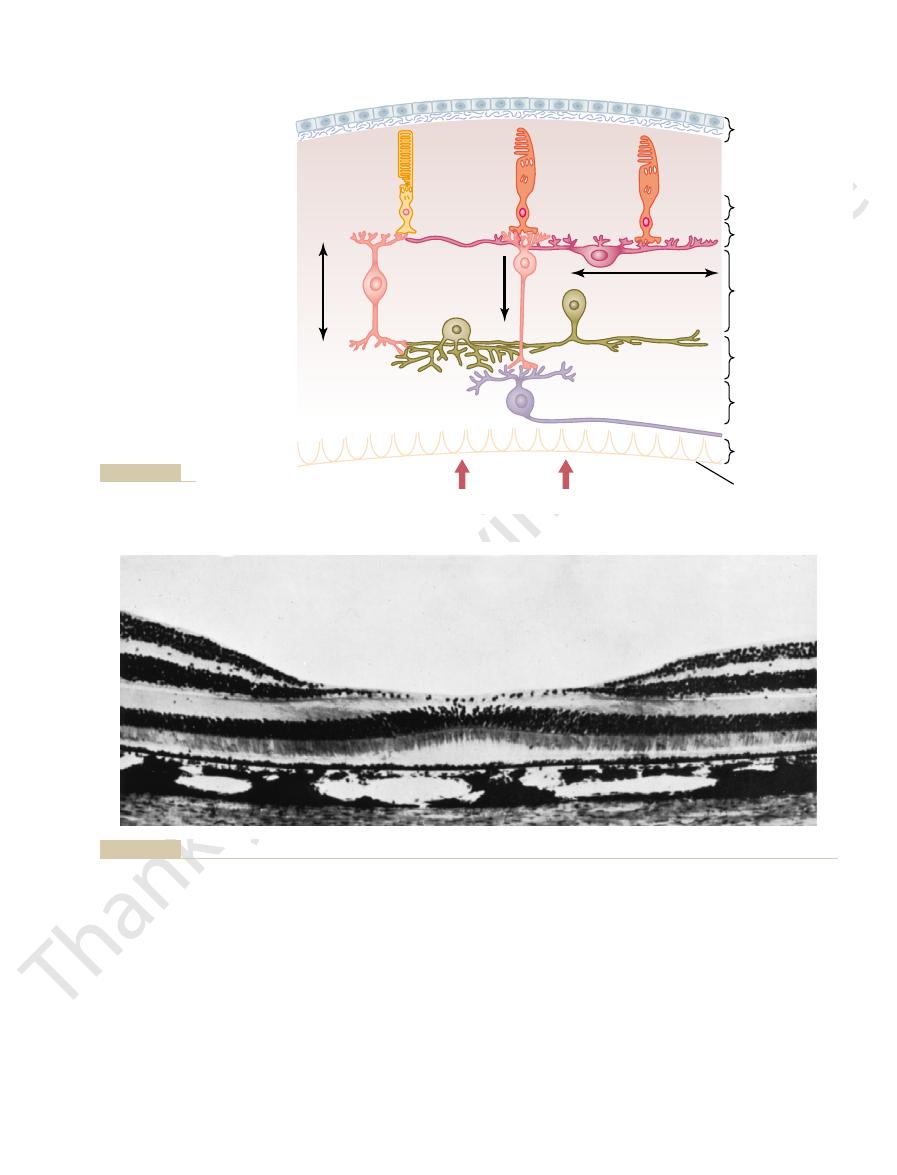
(see Figure 50–1); that is, it passes first
ous humor, it
fibers, and (9) inner limiting membrane.
nuclear layer, (6) inner plexiform layer, (7) ganglionic layer, (8) layer of optic nerve
taining the cell bodies of the rods and cones, (4) outer plexiform layer, (5) inner
(2) layer of rods and cones projecting to the pigment, (3) outer nuclear layer con-
are arranged in layers from the outside to the inside as follows: (1) pigmented layer,
Figure 50–1 shows the functional components of the retina which
signals.
optic nerve fibers and the cerebral cortex. The
layers of neurons in the retina itself and, finally, into
the dark. When either rods or cones are excited,
rods,
for color vision, and (2) the
cones,
The retina is the light-sensitive portion of the eye
The Eye: II. Receptor and
C
H
A
P
T
E
R
5
0
626
Neural Function of the Retina
that contains (1) the
which are responsible
which are mainly
responsible for black and white vision and vision in
signals are transmitted first through successive
purpose of this chapter is to explain the mechanisms by which the rods and
cones detect light and color and convert the visual image into optic nerve
Anatomy and Function of the Structural
Elements of the Retina
Layers of the Retina.
After light passes through the lens system of the eye and then through the vitre-
enters the retina from the inside
through the ganglion cells and then through the plexiform and nuclear layers before
it finally reaches the layer of rods and cones located all the way on the outer edge
Foveal Region of the Retina and Its Importance in Acute Vision.
is a minute area
these cones have a special structure that aids their detection of detail in the visual
and plexiform layers are all displaced to one side rather than resting directly on top
Rods and Cones.
outer segment, (2) the inner segment, (3) the
and (4)
the
rhodopsin;
The Eye: II. Receptor and Neural Function of the Retina
Chapter 50
627
Pigmented layer
Cone
Cone
Vertical
pathway
Lateral pathway
Rod
Outer nuclear layer
Horizontal cell
Amacrine cell
Bipolar
cell
Bipolar
cell
Amacrine
cell
Distal
Proximal
Inner nuclear layer
Inner plexiform
layer
Ganglion cell
To optic nerve
Ganglion cell layer
Stratum opticum
Inner limiting
membrane
Outer plexiform
layer
DIRECTION OF LIGHT
Layers of retina.
Figure 50–1
that connects with subsequent neuronal cells, the
The
ing energy for function of the photoreceptors.
larly important are the mitochondria; as explained later,
usual cytoplasm with cytoplasmic organelles. Particu-
The
proteins. The concentrations of these photosensitive
gated proteins. They are incorporated into the mem-
cone.
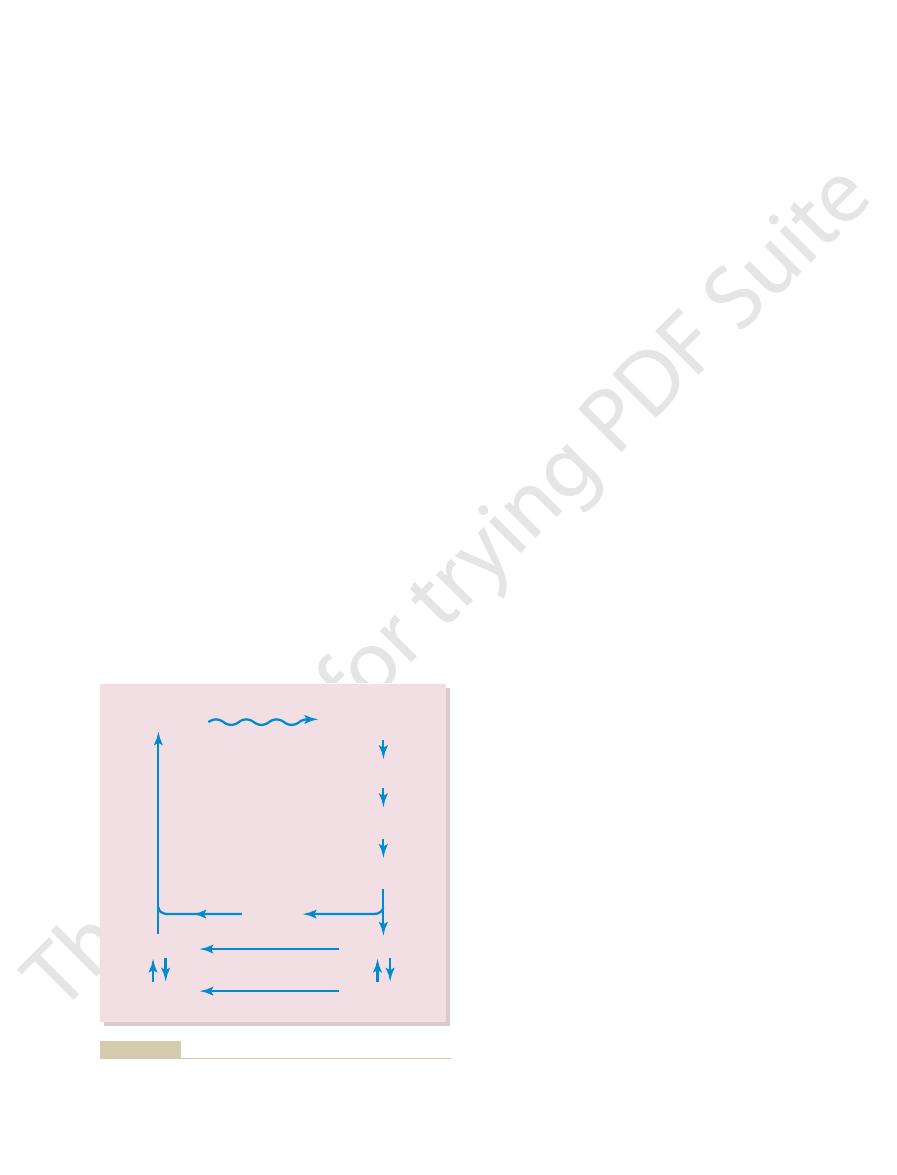
brane. There are as many as 1000 discs in each rod or
discs.
Figures 50–3 and 50–4 the large numbers of
except for differences in spectral sensitivity.
color pigments,
photochemicals, usually called simply
1986; courtesy H. Mizoguchi.)
interference with light transmission. (From Fawcett DW: Bloom and Fawcett: A Textbook of Histology, 11th ed. Philadelphia: WB S
o decrease
Photomicrograph of the macula and of the fovea in its center. Note that the inner layers of the retina are pulled to the side t
Figure 50–2
aunders,
that function almost exactly the same as rhodopsin
Note in the outer segments of the rods and cones in
Each
of the discs is actually an infolded shelf of cell mem-
Both rhodopsin and the color pigments are conju-
branes of the discs in the form of transmembrane
pigments in the discs are so great that the pigments
themselves constitute about 40 per cent of the entire
mass of the outer segment.
inner segment of the rod or cone contains the
these mitochondria play the important role of provid-
synaptic body is the portion of the rod or cone
color pigments,
cones,
nerve fibers leading from the eye. The light-sensitive
pose on exposure to light and, in the process, excite the
Photochemistry of Vision
will be unable to function even after surgical repair.
replaced soon, however, the retina will be destroyed and
normal relation with the pigment epithelium. If it is not
to the neural retina through the retinal artery, the
toward the interior of the globe.
in the vitreous humor, which pull areas of the retina
retina and the pigment epithelium. Detachment is occa-
. In some instances,
detaches from the pigment epithelium
The neural retina occasionally
blood vessels for their nutrition, especially for their
and cones, depend mainly on diffusion from the choroid
of the retina, especially the outer segments of the rods
lying between the retina and the sclera. The outer layers
choroid,
However, the outermost layer of the retina is adher-
structures of the eye.
Thus, the inner layers of the retina
inside retinal surface.
artery, which enters the eyeball through the center of
The nutrient blood supply for the internal
Blood Supply of the Retina—The Central Retinal Artery and the
rods and cones.
the pigment. We show later that vitamin A is an impor-
the rods and cones, which themselves are embedded in
This vitamin A is exchanged back and forth
The pigment layer also stores large quantities of
normal 20/20 values.
of albinos, even with the best optical correction, is
and excites many receptors. Therefore, the visual acuity
the retina and by the underlying sclera, so that a
their bodies. When an albino enters a bright room, light
albinos,
The importance of melanin in the pigment layer is
precise images.
camera. Without it, light rays would be reflected in all
clear vision. This pigment performs the same function
the globe of the eyeball; this is extremely important for
The black pigment
, that represent the next
horizontal
The Nervous System: B. The Special Senses
628
Unit X
and bipolar cells
stages in the vision chain.
Pigment Layer of the Retina.
melanin in
the pigment layer prevents light reflection throughout
in the eye as the black coloring inside the bellows of a
directions within the eyeball and would cause diffuse
lighting of the retina rather than the normal contrast
between dark and light spots required for formation of
well illustrated by its absence in
people who are
hereditarily lacking in melanin pigment in all parts of
that impinges on the retina is reflected in all directions
inside the eyeball by the unpigmented surfaces of
single discrete spot of light that would normally
excite only a few rods or cones is reflected everywhere
seldom better than 20/100 to 20/200 rather than the
vitamin A.
through the cell membranes of the outer segments of
tant precursor of the photosensitive chemicals of the
Choroid.
layers of the retina is derived from the central retinal
the optic nerve and then divides to supply the entire
have their own blood supply independent of the other
ent to the
which is also a highly vascular tissue
oxygen.
Retinal Detachment.
the cause of such detachment is injury to the eyeball
that allows fluid or blood to collect between the neural
sionally caused by contracture of fine collagenous fibrils
Partly because of diffusion across the detachment gap
and partly because of the independent blood supply
detached retina can resist degeneration for days and can
become functional again if it is surgically replaced in its
Both rods and cones contain chemicals that decom-
chemical in the rods is called rhodopsin; the light-
sensitive chemicals in the
called cone pigments
or
have compositions only slightly dif-
ferent from that of rhodopsin.
Outer segment
Membrane shelves
lined with rhodopsin
or color pigment
Mitochondria
Inner segment
Nucleus
Outer limiting
membrane
Synaptic body
Schematic drawing of the functional parts of the rods and cones.
Figure 50–3
(Courtesy Dr. Richard Young.)
Membranous structures of the outer segments of a rod
Figure 50–4
(left) and
a cone (right).
When the rod is exposed to light, the resulting
Once night blindness develops, it can sometimes be
stored in the liver and can be made available to the eyes.
because large quantities of vitamin A are normally
remain on a vitamin A–deficient diet for months,
For night blindness to occur, a person usually must
adequate vision in vitamin A–deficient persons.
This condition is called
this is that without vitamin A, the amounts of retinal and
with severe vitamin A deficiency. The simple reason for
light intensities.
sion between retinal and vitamin A is especially impor-
in the retina. We shall see later that this interconver-
A, thus reducing the amount of light-sensitive pigment
retinal in the retina, it is converted back into vitamin
retinal when needed. Conversely, when there is excess
vitamin A is normally always available to form new
rods and in the pigment layer of the retina. Therefore,
Vitamin A is present both in the cytoplasm of the
retinal, which combines with scotopsin to form new
Finally, the 11-
retinol under the influence of the enzyme isomerase.
Then the all-
retinol, which is one form of vitamin A.
into all-
retinal. This is by conversion of the all-
Figure 50–5 that there is a second chemical route by
light energy.
to re-form rhodopsin, which then remains stable until
formed, it automatically recombines with the scotopsin
retinal isomerase.
retinal. This process
of rhodopsin, as shown in Figure 50–5, is to reconvert
The first stage in re-formation
action potential, as we discuss later.
that excites electrical changes in the rods,
It is the metarhodopsin II, also called
seconds), into the completely split products
and finally, much more slowly (in
This then decays in microseconds
retinal and scotopsin. Bathorhodopsin
away from the scotopsin. The immediate product is
a straight molecule rather than an angulated molecule.
in the retinal portion of the rhodopsin, which leads to
50–5. The cause of this is photoactivation of electrons
fraction of a second, as shown at the top of Figure
When light energy is absorbed by rhodopsin, the
retinal. This
(also called “retinene”). Furthermore, the retinal is a
This substance is a combination of the
visual purple.
The outer
Rhodopsin and Its Decomposition by Light Energy.
Excitation of the Rods
Rhodopsin-Retinal Visual Cycle, and
applied to the cone pigments.
chemistry of rhodopsin, but the same principles can be
In this section, we discuss principally the photo-
The Eye: II. Receptor and Neural Function of the Retina
Chapter 50
629
segment of the rod that projects into the pigment layer
of the retina has a concentration of about 40 per cent
of the light-sensitive pigment called rhodopsin, or
protein scotopsin and the carotenoid pigment retinal
particular type called 11-cis
cis form of
retinal is important because only this form can bind
with scotopsin to synthesize rhodopsin.
rhodopsin begins to decompose within a very small
instantaneous change of the cis form of retinal into an
all-trans form that still has the same chemical structure
as the cis form but has a different physical structure—
Because the three-dimensional orientation of the
reactive sites of the all-trans retinal no longer fits
with the orientation of the reactive sites on the protein
scotopsin,
the all-trans
retinal begins to pull
bathorhodopsin, which is a partially split combination
of the all-trans
is extremely unstable and decays in nanoseconds to
lumirhodopsin.
to metarhodopsin I, then in about a millisecond to
metarhodopsin II,
scotopsin
and all-trans retinal.
activated
rhodopsin,
and the rods then transmit the visual image into the
central nervous system in the form of optic nerve
Re-formation of Rhodopsin.
the all-trans retinal into 11-cis
requires metabolic energy and is catalyzed by the
enzyme
Once the 11-cis retinal is
its decomposition is again triggered by absorption of
Role of Vitamin A for Formation of Rhodopsin.
Note in
which all-trans retinal can be converted into 11-cis
trans retinal first
trans
trans retinol is converted into 11-cis
cis retinol is converted into 11-cis
rhodopsin.
tant in long-term adaptation of the retina to different
Night Blindness.
Night blindness occurs in any person
rhodopsin that can be formed are severely depressed.
night blindness because the
amount of light available at night is too little to permit
reversed in less than 1 hour by intravenous injection of
vitamin A.
Excitation of the Rod When Rhodopsin
Is Activated by Light
The Rod Receptor Potential Is Hyperpolarizing, Not Depolariz-
ing.
Rhodopsin
Scotopsin
11-cis retinal
11-cis retinol
all-trans retinal
all-trans retinol
(Vitamin A)
Bathorhodopsin
(nsec)
Lumirhodopsin
(
µ
sec)
Metarhodopsin I
(msec)
Metarhodopsin II
(sec)
Light energy
Isomerase
Isomerase
(p sec)
(minutes)
formation of rhodopsin by the chemical processes.
of rhodopsin during exposure to light and subsequent slow re-
Rhodopsin-retinal visual cycle in the rod, showing decomposition
Figure 50–5
phosphodiesterase.
3. The
2. The
shown in Figure 50–5.
form of rhodopsin, as already discussed and
retinal portion of the rhodopsin; this leads to the
1. The
excitation? The answer is that the photoreceptors
of light will cause half saturation of the rod. How
potential in a rod of about 1 millivolt. Only 30 photons
unit of light energy, can cause a measurable receptor
a single photon of light, the smallest possible quantal
Under optimal conditions,
Mechanism by Which Rhodopsin Decomposition
would be possible otherwise.
of the light intensity. This is exceedingly important,
as in the rods. A visual image impinged on the rods
second. In cones, the change occurs four times as fast
that is, the
When a sudden pulse of light strikes the retina, the
Relation of the Receptor Potential to Light Intensity.
Duration of the Receptor Potential, and Logarithmic
potassium ions across the membrane.
millivolts, which is near the equilibrium potential for
sity, the membrane potential approaches –70 to –80
tronegativity becomes—that is, the greater is the
light energy striking the rod, the greater the elec-
inside the membrane, and the greater the amount of
than leak back in. Because they are positive ions, their
segment. Thus, more sodium ions now leave the rod
the rod, even though sodium ions continue to be
to decompose, and this
of the rod is exposed to light, the rhodopsin begins
Then, when the rhodopsin in the outer segment
receptors.
of the rod, measuring about –40 millivolts rather than
normal dark conditions, when the rod is not excited,
negativity on the inside of the entire cell. Thus,
is very leaky to sodium ions. Therefore, positively
state,
different; here, the rod membrane, in the
where the photoreceptor discs are located, is entirely
the entire cell. However, the outer segment of the rod,
pumps sodium from inside the rod to the outside,
segments of the rod. The inner segment continually
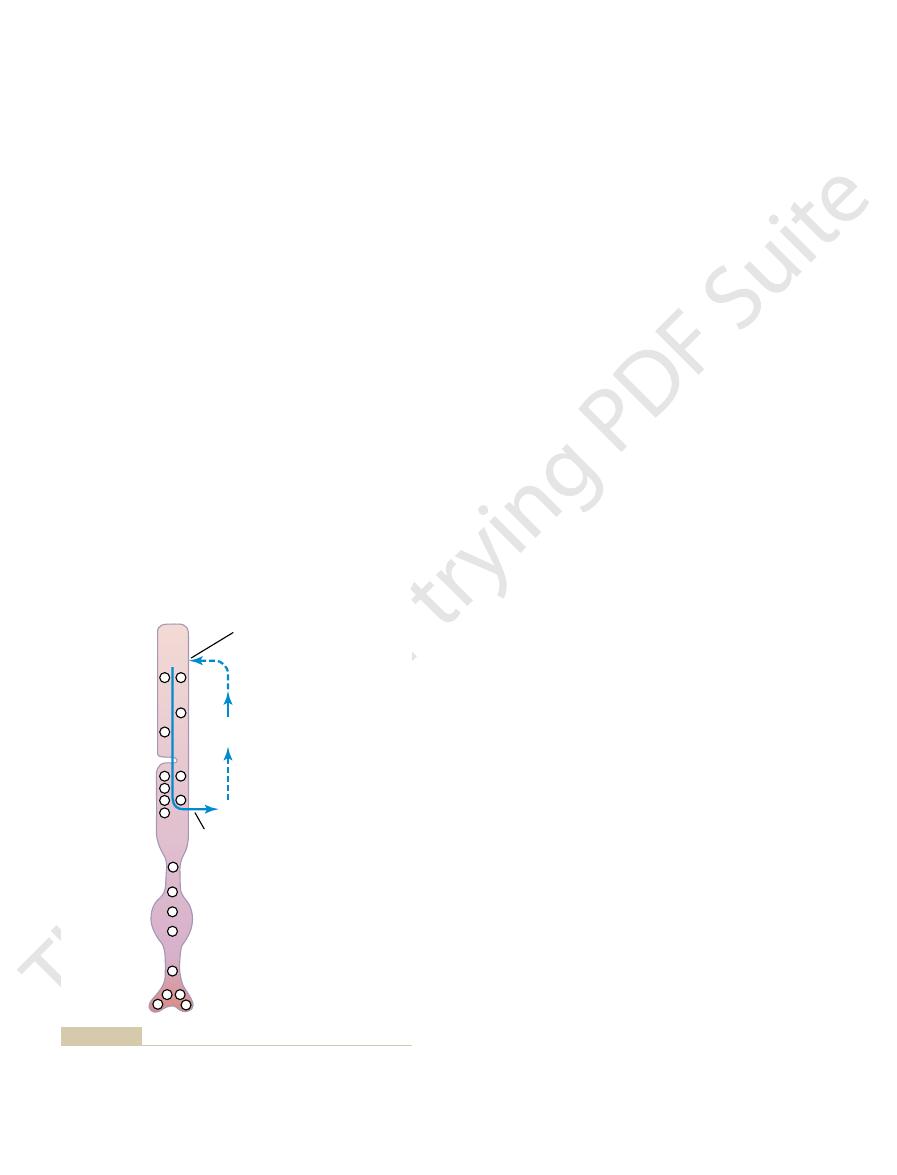
Figure 50–6 shows movement of sodium ions in a
brane in the following way.
This causes hyperpolarization of the entire rod mem-
decomposes, it decreases the rod membrane conduc-
polarization? The answer is that
sensory receptors.
of “depolarization”) that occurs in almost all other
the rod membrane. This is exactly
intrarod membrane potential, which is a state of
tials in almost all other sensory receptors. That is, exci-
The Nervous System: B. The Special Senses
630
Unit X
receptor potential is different from the receptor poten-
tation of the rod causes increased negativity of the
hyper-
polarization, meaning that there is more negativity
than normal inside
opposite to the decreased negativity (the process
But how does activation of rhodopsin cause hyper-
when rhodopsin
tance for sodium ions in the outer segment of the rod.
complete electrical circuit through the inner and outer
thereby creating a negative potential on the inside of
dark
charged sodium ions continually leak back to the
inside of the rod and thereby neutralize much of the
under
there is reduced electronegativity inside the membrane
the usual –70 to –80 millivolts found in most sensory
decreases the outer segment
membrane conductance of sodium to the interior of
pumped outward through the membrane of the inner
loss from inside the rod creates increased negativity
degree of hyperpolarization. At maximum light inten-
transient hyperpolarization that occurs in the rods—
receptor potential that occurs—reaches a
peak in about 0.3 second and lasts for more than a
of the retina for only one millionth of a second can
sometimes cause the sensation of seeing the image for
longer than a second.
Another characteristic of the receptor potential is
that it is approximately proportional to the logarithm
because it allows the eye to discriminate light intensi-
ties through a range many thousand times as great as
Decreases
Membrane Sodium Conductance—The
Excitation “Cascade.”
can such a small amount of light cause such great
have an extremely sensitive chemical cascade that
amplifies the stimulatory effects about a millionfold,
as follows:
photon activates an electron in the 11-cis
formation of metarhodopsin II, which is the active
activated rhodopsin functions as an enzyme to
activate many molecules of transducin, a protein
present in an inactive form in the membranes of
the discs and cell membrane of the rod.
activated transducin activates many more
molecules of
-
-
- -
-
-
-
-
-
-
-
-
-
- -
-
-
-
-
Leakage current
decreased by
decomposing
rhodopsin
Sodium
current
Sodium
pump
Na
+
Na
+
of the rod.
decreases
Theoretical basis for generation of a “hyperpolarization receptor
Figure 50–6
potential” caused by rhodopsin decomposition, which
the flow of positively charged sodium ions into the outer segment

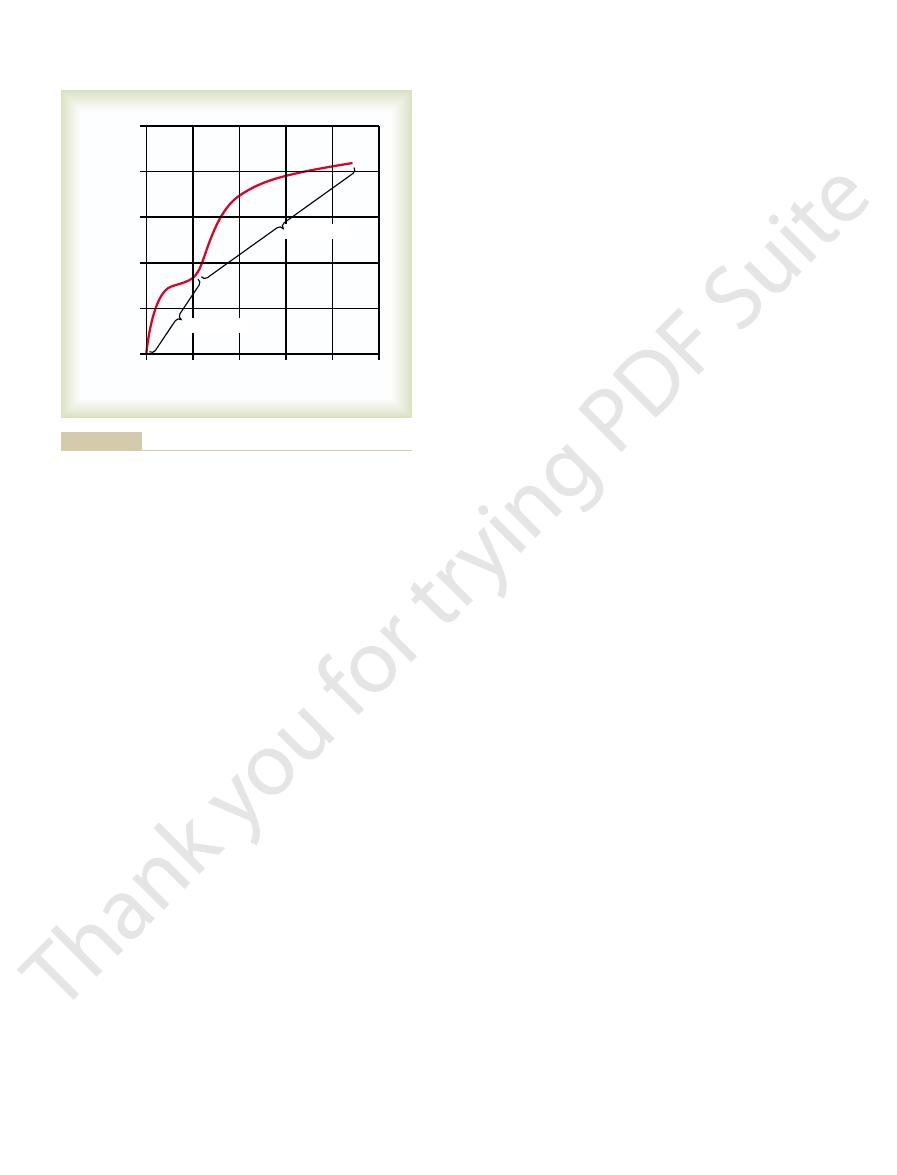
40 minutes, about 25,000-fold.
tivity has increased about 6000-fold, and at the end of
required intensity. At the end of 20 minutes, the sensi-
sensitivity has already increased 10-fold—that is, the
first entering the darkness, but within 1 minute, the
having been exposed to bright light for several hours.
Figure 50–8 shows the course of dark adaptation
rods and cones to combine with the retinal. This is
to give still more light-sensitive pigments, the final
Furthermore, vitamin A is converted back into retinal
are converted back into the light-sensitive pigments.
long time, the retinal and opsins in the rods and cones
Conversely, if a person remains in darkness for a
reduced. This is called
in the rods and cones are considerably reduced, and
into vitamin A. Because of these two effects, the con-
retinal and opsins. Furthermore, much of the retinal of
light for hours, large portions of the photochemicals in
Retinal Sensitivity—Light and
Automatic Regulation of
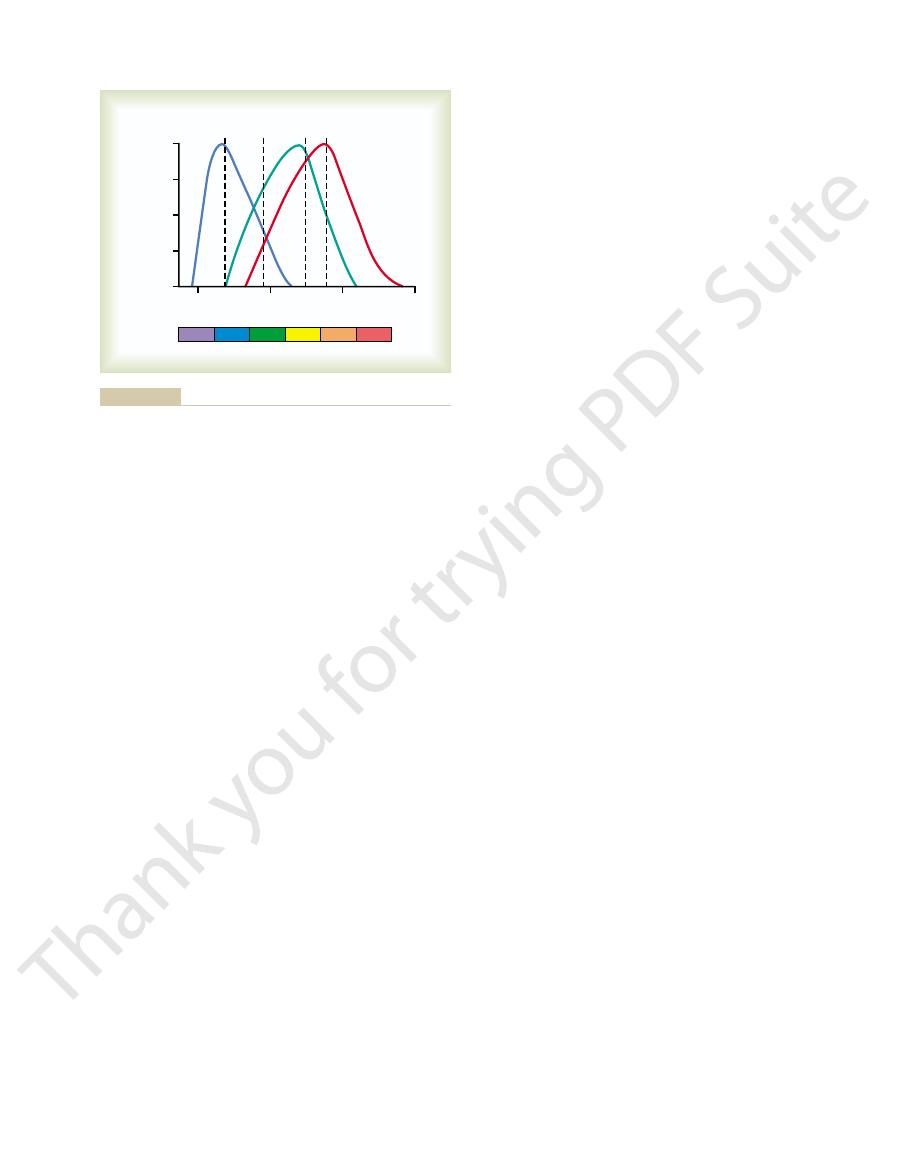
nanometers.
for the rhodopsin of the rods, with a peak at 505
in Figure 50–7. Also shown is the absorption curve
the retina differentiates the colors. The approximate
ity for each type of cone, which begins to explain how
These are also the wavelengths for peak light sensitiv-
lengths of 445, 535, and 570 nanometers, respectively.
The
blue-sensitive pigment, green-
are called, respectively,
colors: blue, green, or red. These color pigments
color pigments is present in each of the different cones,
In the discussion of color vision later in the chapter,
nations of retinal and photopsins.
sensitive pigments of the cones, therefore, are combi-
the same in the cones as in the rods. The color-
The
are slightly different from the scotopsin of the rods.
tions, or the opsins—called
in the rods. The only difference is that the protein por-
Photochemistry of Color Vision by the Cones
than the rods, but even this allows color vision at
The cones are about 30 to 300 times less sensitive
dark conditions.
This explains the extreme sensitivity of the rods under
of light to cause movement of millions of sodium ions.
Thus, the rods have developed an important chemi-
sodium channels.
(the metarhodopsin II), and the entire cascade
the rod, inactivates the activated rhodopsin
rhodopsin kinase,
5. Within about a second, another enzyme,
ion flow is what excites the rod, as already
channel opens again. This diminution of sodium
rhodopsin. Because the sodium flux through
channels to close. Several hundred channels
phosphodiesterase hydrolyzes the cGMP, this
“splints” it in the open state. But in light, when
of the rod’s outer membrane in a way that
destroying it. Before being destroyed, the cGMP
The Eye: II. Receptor and Neural Function of the Retina
Chapter 50
631
4. Activated phosphodiesterase is another enzyme;
it immediately hydrolyzes many molecules of
cyclic guanosine monophosphate (cGMP), thus
had been bound with the sodium channel protein
removes the splinting and allows the sodium
close for each originally activated molecule of
each of these channels has been extremely rapid,
flow of more than a million sodium ions is
blocked by the channel closure before the
discussed.
which is always present in
reverses back to the normal state with open
cal cascade that amplifies the effect of a single photon
any intensity of light greater than extremely dim
twilight.
It was pointed out at the outset of this discussion that
the photochemicals in the cones have almost exactly
the same chemical composition as that of rhodopsin
photopsins in the cones—
retinal portion of all the visual pigments is exactly
it will become evident that only one of three types of
thus making the cones selectively sensitive to different
sensitive pigment, and red-sensitive pigment.
absorption characteristics of the pigments in the three
types of cones show peak absorbencies at light wave-
absorption curves for these three pigments are shown
Dark Adaptation
Light and Dark Adaptation.
If a person has been in bright
both the rods and the cones will have been reduced to
both the rods and the cones will have been converted
centrations of the photosensitive chemicals remaining
the sensitivity of the eye to light is correspondingly
light adaptation.
limit being determined by the amount of opsins in the
called dark adaptation.
when a person is exposed to total darkness after
Note that the sensitivity of the retina is very low on
retina can respond to light of one tenth the previously
600
700
Yellow
Violet
Blue
Green
400
Blue
cone
Green
cone
Red
cone
Rods
500
Red
Orange
Wavelength (nanometers)
Light absorption
(per cent of maximum)
0
25
50
75
100
and cones of the human retina: direct measurements reveal mech-
1964, and by Brown PK, Wald G: Visual pigments in single rods
Jr: Visual pigments of single primate cones. Science 143:1181,
from curves recorded by Marks WB, Dobelle WH, MacNichol EF
of the three color-receptive cones of the human retina. (Drawn
Light absorption by the pigment of the rods and by the pigments
Figure 50–7
anisms of human night and color vision. Science 144:45, 1964.
“ 1964 by the American Association for the Advancement of
Science.)
ring to Figure 50–9, one can see that an orange
Refer-
Interpretation of Color in the Nervous System.
ently in Figure 50–9. They can explain most of the phe-
curves are shown in Figure 50–7 and slightly differ-
the three types of pigment found in the cones. These
basis of color vision tests, the spectral sensitivities of
Spectral Sensitivities of the Three Types of Cones.
mixed in different combinations.
almost all gradations of colors when only red, green,
Tricolor Mechanism of Color
This section is a discussion of the mechanisms by
From the preceding sections, we have learned that dif-
Color Vision
times that of starlight, yet the eye can function both in
adaptation, the intensity of sunlight is about 10 billion
ter. As an example of the extremes of light and dark
After dark adaptation, the light spots begin to regis-
Conversely, when a person first enters darkness, the
receptors excessively.
its different parts. This is poor vision, and it remains
visual image is bleached, having little contrast among
exceedingly bright, and as a consequence, the entire
sunlight. Then, even the dark spots in the images seem
lighter areas but not to the darker areas. An example
image, it is essential that the sensitivity of the retina
as much as 500,000 to 1 million times, the sensitivity
adaptation, the eye can change its sensitivity to light
Value of Light and Dark Adaptation in Vision.
adaptation by the photochemicals.
neural adaptation occurs in a fraction of a second, in
the neural circuit. Although the degree of adaptation is
glion cells are all intense. However, most of these signals
bipolar cells, horizontal cells, amacrine cells, and gan-
intensity first increases, the signals transmitted by the
in the retina itself and in the brain. That is, when light
The other mechanism is
pupillary opening.
mately 30-fold within a fraction of a second, because of
in Chapter 49. This can cause adaptation of approxi-
change in pupillary size,
other mechanisms for light and dark adaptation. The
rhodopsin or color photochemicals, the eye has two
sensitivity, as discussed later in the chapter.
cell in the retina; these rods summate to increase their
sitivity increasing tremendously. In addition, still more
to adapt for many minutes and even hours, their sen-
a few minutes, while the slowly adapting rods continue
rapid adaptation, the cones cease adapting after only
change in darkness as the rods do. Therefore, despite
rapidly in cones as in rods. However, the cones do not
vision, including adaptation, occur about four times as
tation of the cones, because all the chemical events of
curve. The early portion of the curve is caused by adap-
Note, however, the inflection in the
adaptation curve.
The resulting curve of Figure 50–8 is called the
The Nervous System: B. The Special Senses
632
Unit X
dark
achieve anywhere near the same degree of sensitivity
sensitivity of the rods is caused by neuronal signal con-
vergence of 100 or more rods onto a single ganglion
Other Mechanisms of Light and Dark Adaptation.
In addition
to adaptation caused by changes in concentrations of
first of these is a
as discussed
changes in the amount of light allowed through the
neural adaptation, involving
the neurons in the successive stages of the visual chain
decrease rapidly at different stages of transmission in
only a fewfold rather than the many thousandfold that
occurs during adaptation of the photochemical system,
contrast to the many minutes to hours required for full
Between the
limits of maximal dark adaptation and maximal light
automatically adjusting to changes in illumination.
Because registration of images by the retina
requires detection of both dark and light spots in the
always be adjusted so that the receptors respond to the
of maladjustment of retinal adaptation occurs when a
person leaves a movie theater and enters the bright
poor until the retina has adapted sufficiently so that
the darker areas of the image no longer stimulate the
sensitivity of the retina is usually so slight that even
the light spots in the image cannot excite the retina.
bright sunlight after light adaptation and in starlight
after dark adaptation.
ferent cones are sensitive to different colors of light.
which the retina detects the different gradations of
color in the visual spectrum.
Detection
All theories of color vision are based on the well-
known observation that the human eye can detect
and blue monochromatic lights are appropriately
On the
the three types of cones in humans have proved to be
essentially the same as the light absorption curves for
nomena of color vision.
20
30
40
50
0
10
Minutes in dark
Retinal sensitivity
1
10
100
1000
10,000
100,000
Rod adaptation
Cone adaptation
rod adaptation.
Dark adaptation, demonstrating the relation of cone adaptation to
Figure 50–8
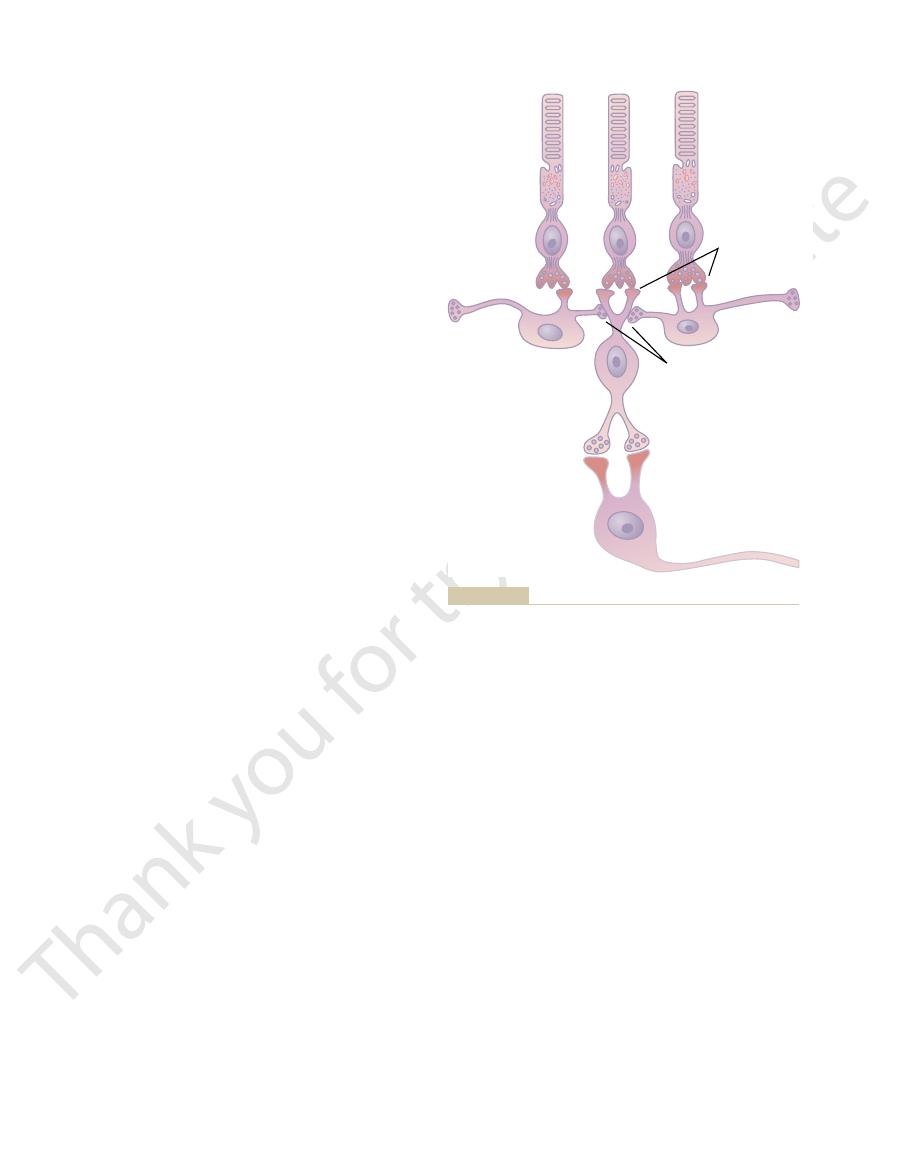
inner plexiform layer, where they synapse with
from the rods, cones, and horizontal cells to the
bipolar cells,
3. The
horizontal cells,
2. The
plexiform layer, where they synapse with bipolar
1. The photoreceptors themselves—the
foveal retina. The different neuronal cell types are as
neural connections, showing at the left the circuit in
Figure 50–11 presents the essentials of the retina’s
neural organization in the retina. To simplify this,
Figure 50–1 shows the tremendous complexity of
Neural Circuitry of the Retina
certain colors by color-blind people.
cones depicted in Figure 50–9, it can be readily under-
“2,” and the green-blind person reads “4.”
vision reads “42,” whereas the red-blind person reads
“21.” In the bottom chart, the person with normal color
“74,” whereas the red-green color-blind person reads
top chart, the person with normal color vision reads
a confusion of spots of several different colors. In the
shown in Figure 50–10. These charts are arranged with
Color Test Charts.
nomenon called blue weakness.
although sometimes they are underrepresented, which
Only rarely are blue cones missing,
Blue Weakness.
blindness is passed from mother to son, and the mother
inherited from the mother, never from the father, color
can lead to color blindness.
the male has only one X chromosome, a missing gene
always has a normal gene for each type of cone. Because
Yet color blindness almost never occurs in females
female X chromosome code for the respective cones.
occurs almost exclusively in males. That is, genes in the
detect the long wavelength red color.
cones. A color-blind person who lacks green cones is
blindness.
colors; the person is especially unable to distinguish red
cones. If either of these two cones is missing, the person
wavelengths of 525 and 675 nanometers, are normally
orange, and red colors, which are the colors between the
instance, one can see in Figure 50–9 that green, yellow,
unable to distinguish some colors from others. For
receptive cones is missing from the eye, the person is
When a single group of color-
types of cones about equally.
thermore, the perception of white can be achieved by
bination of all the wavelengths of the spectrum. Fur-
of light corresponding to white; instead, white is a com-
tion of seeing white. Yet there is no single wavelength
all the red, green, and blue cones gives one the sensa-
preted as yellow, and 31:67:36 as green.
system as blue. Likewise, ratios of 83:83:0 are inter-
value of 0, and the blue cones to a value of 97. This set
cones to a stimulus value of 0, the green cones to a
orange. Conversely, a monochromatic blue light with a
of cones in this instance are 99:42:0. The nervous
at all. Thus, the ratios of stimulation of the three types
to a stimulus value of about 42, but the blue cones not
at optimum wavelength); it stimulates the green cones
The Eye: II. Receptor and Neural Function of the Retina
Chapter 50
633
monochromatic light with a wavelength of 580
nanometers stimulates the red cones to a stimulus
value of about 99 (99 per cent of the peak stimulation
system interprets this set of ratios as the sensation of
wavelength of 450 nanometers stimulates the red
of ratios—0:0:97—is interpreted by the nervous
Perception of White Light.
About equal stimulation of
stimulating the retina with a proper combination of
only three chosen colors that stimulate the respective
Color Blindness
Red-Green Color Blindness.
distinguished from one another by the red and green
cannot use this mechanism for distinguishing these four
from green and is therefore said to have red-green color
A person with loss of red cones is called a protanope;
the overall visual spectrum is noticeably shortened at
the long wavelength end because of a lack of the red
called a deuteranope; this person has a perfectly normal
visual spectral width because red cones are available to
Red-green color blindness is a genetic disorder that
because at least one of the two X chromosomes almost
Because the X chromosome in the male is always
is said to be a color blindness carrier; this is true in about
8 per cent of all women.
is a genetically inherited state giving rise to the phe-
A rapid method for determining color
blindness is based on the use of spot charts such as those
If one studies these charts while at the same time
observing the spectral sensitivity curves of the different
stood how excessive emphasis can be placed on spots of
Neural Function of the Retina
the peripheral retina and at the right the circuit in the
follows:
rods and
cones—which transmit signals to the outer
cells and horizontal cells
which transmit signals
horizontally in the outer plexiform layer from the
rods and cones to bipolar cells
which transmit signals vertically
ganglion cells and amacrine cells
97
99
600
700
Yellow
Violet
Blue
Green
400
Blue
cone
Green
cone
Red
cone
500
Red
Orange
Wavelength (nanometers)
Light absorption
(per cent of maximum)
Blue
Green
Yellow
Orange
0
25
50
67
36
42
83
83
31
0
75
100
green, yellow, and orange.
sensitive cones by monochromatic lights of four colors: blue,
Demonstration of the degree of stimulation of the different color-
Figure 50–9

trast in the visual image.
Their role may be to help control the degree of con-
by the horizontal cells in the outer plexiform layer.
outer plexiform layer. These signals are inhibitory and
cell. This cell transmits signals in the retro-
prominent and not shown in the figure, is the
A sixth type of neuronal cell in the retina, not very
ganglion cells,
5. The
directions, either directly from bipolar cells to
amacrine cells,
4. The
The Nervous System: B. The Special Senses
634
Unit X
which transmit signals in two
ganglion cells or horizontally within the inner
plexiform layer from axons of the bipolar cells to
dendrites of the ganglion cells or to other
amacrine cells
which transmit output signals
from the retina through the optic nerve into the
brain
inter-
plexiform
grade direction from the inner plexiform layer to the
are believed to control lateral spread of visual signals
for Colour Blindness. Tokyo: Kanehara & Co., but tests for
normal person reads “42.” (Reproduced from Ishihara’s Tests
“2,” but the green-blind person (deuteranope) reads “4.” The
In this chart, the red-blind person (protanope) reads
reads “74,” but the red-green color-blind person reads “21.”
In this chart, the normal person
Two Ishihara charts.
Figure 50–10
Upper:
Lower:
color blindness cannot be conducted with this material. For
accurate testing, the original plates should be used.)

light focused on the retina. The visual pathway from
strated in Figure 50–12, which shows a minute spot of
proper visual contrast. This phenomenon is demon-
that is important in all other sensory systems—that is,
Therefore, this lateral connection
always inhibitory.
bipolar cells. The outputs of the horizontal cells
cones, as well as connecting with the dendrites of the
The horizontal cells, shown in Figure 50–11, connect
Lateral Inhibition to Enhance Visual Contrast—
illumination; the signal is not all or none, as would be
the rods and cones, the strength of the hyperpolariz-
of signal strength. Thus, for
The importance of electrotonic conduction is that it
tric current flow, not by action potentials.
bipolar cell or horizontal cell, once again the signal is
tic body, and no action potential is required. Then,
current flow in the cytoplasm all the way to the synap-
segment of a rod or a cone, almost the same degree of
is by electrotonic conduction. That is, when hyperpo-
the visual signals are generated, to the synaptic bodies
cones, conduction from their outer segments, where
the way to the output synapses. Even in the rods and
tric current, not action potentials, in the neuronal cyto-
explained as follows.
Otherwise, all the retinal neurons conduct their visual
importance of these action potentials is questionable.
also been recorded in amacrine cells, although the
the optic nerve. Occasionally, action potentials have
means of action potentials are the ganglion cells, and
The only
Transmission of Most Signals Occurs in the Retinal Neurons by
release inhibitory transmitters.
unclear, but at least some of the horizontal cells
bipolar, horizontal, and interplexiform cells are
as inhibitory transmitters. The transmitters of the
indolamine,
choline,
gamma-aminobutyric acid, glycine, dopamine, acetyl-
least eight types of transmitter substances, including
at their synapses with the bipolar cells.
eated. However, both the rods and the cones release
directly to ganglion cells and by way of amacrine cells.
and cones; the outputs of these bipolar cells pass both
retinal circuitry of Figure 50–11 connect with both rods
The other two bipolar cells shown in the peripheral
connectivity.
Also, horizontal and amacrine cells provide lateral
cells, (3) amacrine cells, and (4) ganglion cells.
in the direct visual pathway: (1) rods, (2) bipolar
cells. Thus, for pure rod vision, there are four neurons
amacrine cells, which relay the signals to the ganglion
The output from the bipolar cell passes only to
type of visual system present in many lower animals.
middle of these connects only to rods, representing the
cones are present. Three bipolar cells are shown; the
tions for the peripheral retina, where both rods and
To the left in Figure 50–11 are the neural connec-
inner plexiform layer.
inhibitory signals laterally in the outer plexiform layer,
ganglion cells. In addition, horizontal cells transmit
the direct pathway: (1) cones, (2) bipolar cells, and (3)
new, fast cone system. This shows three neurons in
foveal portion of the retina,
To the right in Figure 50–11 is the visual pathway
follows.
the circuitry for the two systems is slightly different, as
ducted to the brain two to five times as rapidly. Also,
visual signals for rod vision, and the signals are con-
of vision based on cone vision. The neurons and nerve
of our other sensory systems, the retina has both an
tions Differently from the Rod Pathway.
The Visual Pathway from the Cones to the Ganglion Cells Func-
The Eye: II. Receptor and Neural Function of the Retina
Chapter 50
635
As is true for many
old type of vision based on rod vision and a new type
fibers that conduct the visual signals for cone vision
are considerably larger than those that conduct the
from the
representing the
and amacrine cells transmit signals laterally in the
Neurotransmitters Released by Retinal Neurons.
Not all the
neurotransmitter chemical substances used for synap-
tic transmission in the retina have been entirely delin-
glutamate
Histological and pharmacological studies have shown
there to be many types of amacrine cells secreting at
and
all of which normally function
Electrotonic Conduction, Not by Action Potentials.
retinal neurons that always transmit visual signals by
they send their signals all the way to the brain through
signals by electrotonic conduction, which can be
Electrotonic conduction means direct flow of elec-
plasm and nerve axons from the point of excitation all
larization occurs in response to light in the outer
hyperpolarization is conducted by direct electric
when the transmitter from a rod or cone stimulates a
transmitted from the input to the output by direct elec-
allows graded conduction
ing output signal is directly related to the intensity of
the case for each action potential.
Function of the Horizontal Cells
laterally between the synaptic bodies of the rods and
are
provides the same phenomenon of lateral inhibition
helping to ensure transmission of visual patterns with
Rods
Rod nuclei
Bipolar
cells
Horizontal
cells
Amacrine
cells
Ganglion
cells
Cones
Pigment layer
area to the right.
Neural organization of the retina: peripheral area to the left, foveal
Figure 50–11

peripherally.
cones, as shown to the right in Figure 50–11. This
cones—about 35,000 of them—and no rods. Also, the
central fovea,
the center, in the
increase the acuity of vision in the central retina. In
become more slender. These effects progressively
on each optic fiber, and the rods and cones also
approaches the fovea, fewer rods and cones converge
peripheral retina and the central retina. As one
However, major differences exist between the
about 1.6 million. Thus, an average of 60 rods and 2
million cones; yet the number of ganglion cells is only
In a sense, then, many or most amacrine cells are
directional sensitive.
direction; therefore, these amacrine cells are said to be
illumination, irrespective of direction.
turned either on or off, signaling simply a change in
of visual signals, but again, the response dies quickly.
dies rapidly.
the onset of a continuing visual signal, but the response
amacrine cells to ganglion cells.
for rod vision—that is, from rod to bipolar cells to
been characterized, and all of them are different. One
by morphological or histochemical means. The func-
ates over a much greater distance.
between two adjacent photoreceptors. In contrast, the
in the visual image, even when the border lies exactly
bipolar cells lie immediately against each other, this
mechanism. Because depolarizing and hyperpolarizing
lateral inhibition, in addition to the horizontal cell
signals. We shall see later that both positive and nega-
bipolar responses, the importance of this phenomenon
polarity of the electrical response.
zontal cell is an inhibitory cell, this would reverse the
indirectly through a horizontal cell. Because the hori-
rods and cones, whereas the other receives its signal
ing by hyperpolarizing. The other possibility is that one
released by the rods and cones, and the other respond-
ence. One explanation is that the two bipolar cells are
There are two possible explanations for this differ-
hyperpolarize.
when the rods and cones are excited, and others
That is, some bipolar cells depolarize
and inhibitory signals in the visual pathway: (1) the
Two types of bipolar cells provide opposing excitatory
borders in the visual image.
allow high visual accuracy in transmitting contrast
inhibition in the surrounding areas. This is essential to
in the plexiform layers, transmission through the hor-
whereas an area to the side is inhibited. In other words,
The Nervous System: B. The Special Senses
636
Unit X
the centralmost area where the light strikes is excited,
instead of the excitatory signal spreading widely in the
retina because of spreading dendritic and axonal trees
izontal cells puts a stop to this by providing lateral
Some of the amacrine cells probably provide addi-
tional lateral inhibition and further enhancement of
visual contrast in the inner plexiform layer of the
retina as well.
Excitation of Some Bipolar Cells and
Inhibition of Others—The Depolarizing
and Hyperpolarizing Bipolar Cells
depolarizing bipolar cell and (2) the hyperpolarizing
bipolar cell.
of entirely different types—one responding by depo-
larizing in response to the glutamate neurotransmitter
of the bipolar cells receives direct excitation from the
Regardless of the mechanism for the two types of
is that it allows half the bipolar cells to transmit
positive signals and the other half to transmit negative
tive signals are used in transmitting visual information
to the brain.
Another important aspect of this reciprocal relation
between depolarizing and hyperpolarizing bipolar
cells is that it provides a second mechanism for
provides a mechanism for separating contrast borders
horizontal cell mechanism for lateral inhibition oper-
Amacrine Cells and Their Functions
About 30 types of amacrine cells have been identified
tions of about half a dozen types of amacrine cells have
type of amacrine cell is part of the direct pathway
Another type of amacrine cell responds strongly at
Other amacrine cells respond strongly at the offset
Still other amacrine cells respond when a light is
Still another type of amacrine cell responds to
movement of a spot across the retina in a specific
interneurons that help analyze visual signals before
they ever leave the retina.
Ganglion Cells and Optic Nerve Fibers
Each retina contains about 100 million rods and 3
cones converge on each ganglion cell and the optic
nerve fiber leading from the ganglion cell to the brain.
there are only slender
number of optic nerve fibers leading from this part of
the retina is almost exactly equal to the number of
explains the high degree of visual acuity in the central
retina in comparison with the much poorer acuity
Light beam
Neither excited
nor inhibited
Excited area
Inhibited area
Excitation and inhibition of a retinal area caused by a small beam
Figure 50–12
of light, demonstrating the principle of lateral inhibition.

change
This capability of the eyes to detect
have similar transient responses themselves.
partly generated by the amacrine cells, many of which
depolarizing and hyperpolarizing bipolar cells, and the
these responses to light are caused, respectively, by the
and “off-on” responses. The opposite directions of
effects occur. Thus, these records are called “on-off”
inhibition. Then, when the light is turned off, opposite
located lateral to the spot of light; this cell is markedly
second. The lower tracing is from a ganglion cell
on, but decreasing rapidity in the next fraction of a
Figure 50–13. The upper panel shows rapid impulses
in light intensity. This is
changes
As noted previously, many ganglion cells are
Transmission of Changes in Light Intensity—The On-Off
background ganglion cell firing.
The visual signals, in turn, are superimposed onto this
pulses at rates varying between 5 and 40 per second.
when unstimulated, they still transmit continuous im-
itive action potentials instead. Furthermore, even
within the retina is no longer appropriate; therefore,
tion employed in the rods, cones, and bipolar cells
distance involved, the electrotonic method of conduc-
of the optic nerve lead into the brain. Because of the
Excitation of the Ganglion Cells
location of the event, other than to give appropriate
field, but without specifying with great accuracy the
of a second. These ganglion cells presumably apprise
amacrine cells, to rapid changes in the visual image—
The Y ganglion cells respond, like many of the
from widespread retinal areas.
dritic fields, so that signals are picked up by these cells
only 5 per cent of the total. Also, they have broad den-
least numerous of all the ganglion cells, representing
signals to the brain at 50 m/sec or faster. They are the
35 micrometers in diameter, and they transmit their
The Y cells are the largest of all, up to
Function of the Y Cells to Transmit Instantaneous Changes in
cone, X cell transmission is probably responsible for
details of the visual image are transmitted. Also,
fore, it is mainly through the X cells that the fine
their signals represent discrete retinal locations. There-
do not spread widely in the retina. Because of this,
The X cells have small fields because their dendrites
14 m/sec.
medium diameter, between 10 and 15 micrometers,
representing 55 per cent of the total. They are of
most numerous of the ganglion cells are the X cells,
The
Transmission of the Visual Image and Color by the X Cells.
our crude rod vision under dark conditions.
vision, and they are probably important for much of
experiments, the W cells seem to be especially sensi-
areas.
inner plexiform layer, receiving signals from broad
have broad fields in the peripheral retina because the
by way of small bipolar cells and amacrine cells. They
receive most of their excitation from rods, transmitted
slow velocity of only 8 m/sec. These ganglion cells
small, having a diameter less than 10 micrometers, and
stituting about 40 per cent of all the ganglion cells, are
The W cells, con-
Transmission of Rod Vision by the W Cells.
nated W, X, and Y cells. Each of these serves a differ-
There are three distinct types of ganglion cells, desig-
Three Types of Retinal Ganglion Cells and
nerve fibers.
peripheral portions of the retina, so that signals from
times more sensitive to light than cones are, but it is
sitivity of the peripheral retina to weak light. This
The Eye: II. Receptor and Neural Function of the Retina
Chapter 50
637
Another difference between the peripheral and
central portions of the retina is the much greater sen-
results partly from the fact that rods are 30 to 300
further magnified by the fact that as many as 200 rods
converge on a single optic nerve fiber in the more
the rods summate to give even more intense stimula-
tion of the peripheral ganglion cells and their optic
Their Respective Fields
ent function.
they transmit signals in their optic nerve fibers at the
dendrites of the ganglion cells spread widely in the
On the basis of histology as well as physiologic
tive for detecting directional movement in the field of
and transmit signals in their optic nerve fibers at about
because every X cell receives input from at least one
all color vision.
the Visual Image.
either rapid movement or rapid change in light inten-
sity—sending bursts of signals for only small fractions
the central nervous system almost instantaneously
when a new visual event occurs anywhere in the visual
clues that make the eyes move toward the exciting
vision.
Spontaneous, Continuous Action Potentials in the Ganglion
Cells.
It is from the ganglion cells that the long fibers
ganglion cells transmit their signals by means of repet-
Response.
specifically excited by
demonstrated by the records of nerve impulses in
for a fraction of a second when a light is first turned
inhibited when the light is turned on because of lateral
transient nature of the responses is probably at least
in light
intensity is strongly developed in both the peripheral
Excitation
on
1
2
off
Lateral inhibition
Haven, Conn: Yale University Press, 1955.)
physiological Research into the Process of Reception. New
Perception: A Discussion of Aims, Means, and Results of Electro-
(Modified from Granit R: Receptors and Sensory
glion cell in this area is inhibited by the mechanism of
spot of light and (2) an area adjacent to the excited spot; the gan-
Responses of a ganglion cell to light in (1) an area excited by a
Figure 50–13
lateral
inhibition.
ited by the “opponent” color. Therefore, color analysis
begins to differentiate colors. Thus, each color-contrast
The importance of these color-contrast mechanisms
depolarizing bipolar cell, whereas the other color type
the following: One color type of cone excites the gan-
The mechanism of this opposing effect of colors is
yellow colors.
by yellow) on the other hand, giving a reciprocal
The same type of reciprocal effect occurs between
causing inhibition, or vice versa.
and green cones, with red causing excitation and green
type. For instance, this frequently occurs for the red
Conversely, some of the ganglion cells are excited by
“white” signal.
role in the detection of different colors. Instead, it is a
Therefore, the signal from the ganglion cell plays no
ganglion cell, the signal transmitted through the gan-
the red, blue, and green types—stimulate the same
cones or by only a few. When all three types of cones—
Transmission of Color Signals by the
In summary, the mechanism of lateral inhibition
tuate one another.
bipolar cell. Thus, where visual contrasts occur, the
the bipolar cell, and this allows extra excitation of the
unstimulated. Therefore, this cell does not inhibit
cell. The fact that one of the lateral photoreceptors is
two lateral receptors is in the dark. The bright spot of
Figure 50–14, assume that the central photoreceptor is
border occurs in the visual scene. Referring again to
Now, let us examine what happens when a contrast
cell. The two receptors on each side are connected
The central receptor excites a depolarizing bipolar
50–14, which shows at the top three photoreceptors.
ways. One circuit for this is demonstrated in Figure
horizontal cells are mainly inhibitory. Thus, the direct
tory, while the signals transmitted
is neither stimulated nor inhibited. The reason for this
is, when all the photoreceptors are stimulated equally
When flat light is applied to the entire retina—that
occurs.
mitted to the brain, let us explain how this process
borders in the scene. Because this seems to be the
in the Visual Scene—The Role of
Transmission of Signals Depicting Contrasts
detected. Conversely, the same gnat sitting quietly
retina and the central retina. For instance, a minute
The Nervous System: B. The Special Senses
638
Unit X
gnat flying across the field of vision is instantaneously
remains below the threshold of visual detection.
Lateral Inhibition
Many ganglion cells respond mainly to contrast
major means by which the pattern of a scene is trans-
by the incident light—the contrast type of ganglion cell
is that signals transmitted directly from the photore-
ceptors through depolarizing bipolar cells are excita-
laterally through
hyperpolarizing bipolar cells as well as through
excitatory signal through one pathway is likely to be
neutralized by inhibitory signals through lateral path-
to the same bipolar cell through inhibitory horizontal
cells that neutralize the direct excitatory signal if
all three receptors are stimulated simultaneously by
light.
stimulated by a bright spot of light while one of the
light excites the direct pathway through the bipolar
in the dark causes one of the horizontal cells to remain
signals through the direct and lateral pathways accen-
functions in the eye in the same way that it functions
in most other sensory systems—to provide contrast
detection and enhancement.
Ganglion Cells
A single ganglion cell may be stimulated by several
glion cell is the same for any color of the spectrum.
only one color type of cone but inhibited by a second
blue cones on the one hand and a combination
of red and green cones (both of which are excited
excitation-inhibition relation between the blue and
glion cell by the direct excitatory route through a
inhibits the ganglion cell by the indirect inhibitory
route through a hyperpolarizing bipolar cell.
is that they represent a means by which the retina itself
type of ganglion cell is excited by one color but inhib-
begins in the retina and is not entirely a function of
the brain.
Excitation
Inhibition
H
H
B
G
cells, but inhibition from the horizontal cells to the bipolar cell.
synapses between the rods and the bipolar cell and horizontal
and a ganglion cell (G) in the retina, showing excitation at the
Typical arrangement of rods, horizontal cells (H), a bipolar cell (B),
Figure 50–14

related macular degeneration. Arch Ophthalmol 122:598,
Zarbin MA: Current concepts in the pathogenesis of age-
Retin Eye Res 22:683, 2003.
pigment epithelium: genes, mutations, and diseases. Prog
Thompson DA, Gal A: Vitamin A metabolism in the retinal
Trends Neurosci 26:379, 2003.
Taylor WR, Vaney DI: New directions in retinal research.
Physiol Rev 82:875, 2002.
Schwartz EA: Transport-mediated synapses in the retina.
vision defects. Arch Ophthalmol 118:691, 2000.
Neitz M, Neitz J: Molecular genetics of color vision and color
syndromes. Br J Ophthalmol 88:291, 2004.
Michaelides M, Hunt DM, Moore AT: The cone dysfunction
Neurosci 4:877, 2001.
Masland RH: The fundamental plan of the retina. Nat
tion of the vertebrate retina. Prog Brain Res 131:3, 2001.
Kolb H, Nelson R, Ahnelt P, Cuenca N: Cellular organiza-
tion. New York: Springer, 1997.
Hendee WA, Wells PNT: The Perception of Visual Informa-
tion. Curr Biol 13:R775, 2003.
Hardie RC: Phototransduction: shedding light on transloca-
Neurosci 26:181, 2003.
Gegenfurtner KR, Kiper DC: Color vision. Annu Rev
Rev Neurosci 4:563, 2003.
Gegenfurtner KR: Cortical mechanisms of colour vision. Nat
FEBS Lett 528:17, 2002.
rhodopsin: structural implications for retinal disease.
Garriga P, Manyosa J: The eye photoreceptor protein
tion in vertebrate photoreceptors. Physiol Rev 81:117,
Fain GL, Matthews HR, Cornwall MC, Koutalos Y: Adapta-
Neurobiol 13:421, 2003.
diverse cell types and cone-specific circuitry. Curr Opin
Dacey DM, Packer OS: Colour coding in the primate retina:
Calkins DJ: Seeing with S cones. Prog Retin Eye Res 20:255,
Biol 14:R381, 2004.
Burr D, Ross J: Vision: the world through picket fences. Curr
toreceptors. Trends Neurosci 26:314, 2003.
Berson DM: Strange vision: ganglion cells as circadian pho-
Degeneration. St Louis: Mosby, 1999.
Berger JW, Fine SL, Maguire MG: Age-Related Macular
Walter de Gruyter, 1998.
Backharis W, Kliegl R, Werner JS: Color Vision. Berlin:
ferent pigment regeneration pathways. Neuron 36:1, 2002.
Arshavsky V: Like night and day: rods and cones have dif-
The Eye: II. Receptor and Neural Function of the Retina
Chapter 50
639
References
2001.
2001.
2004.